Introduktion
Metan (CH4) er en vigtig drivhusgas. Emissionen fra naturlige miljøer som f.eks. vådområder, oceaner og sedimenter tegner sig for over 70 % af den globale metanemission i atmosfæren (IPCC, 2007). En vurdering af offentliggjorte data afslørede forskellige metanemissionsrater for vådområder, søer, floder, flodmundinger og oceaner (i faldende rækkefølge, Supplerende figur S1). Naturlige vådområder tegner sig alene for 62 % af den biogene CH4-produktion (Kirschke et al., 2013; Nazaries et al., 2013), og emissioner fra vådområder dominerer den mellemårlige variabilitet af metankilderne (Bousquet et al., 2006). I modsætning hertil bidrager det store areal af marine økosystemer kun med ca. 8 % til de naturlige kilder til CH4 (Nazaries et al., 2013). Inden for den globale oceaniske metanemission bidrager flodmundinger med mindre end 10 % (Bange et al., 1994).
Methanogenese, den biologiske dannelse af metan, udføres af methanogene archaea, som primært producerer metan fra H2/CO2, methylgrupper eller acetat under anoxiske forhold (Thauer et al., 2008). Denne reaktion katalyseres af methyl-coenzym M-reduktase (MCR). Genet mcrA, der koder for en underenhed af dette enzym, er en almindeligt anvendt genmarkør i molekylære undersøgelser (Conrad, 2007; Bridgham et al., 2013). Fordelen ved mcrA-genmarkøren er, at den fanger både de fylogenetiske og funktionelle signaturer af methanogener, hvilket giver en høj sekventeringsdybde for denne særlige funktion (Luton et al., 2002; Borrel et al., 2013; Yang et al., 2014). Et stort antal mcrA-sekvenser blev hentet fra en række forskellige naturlige miljøer. Det offentlige mcrA-datasæt gør det muligt at udtrække generelle økologiske mønstre og undersøge de formende miljøgradienter på globale og regionale skalaer. Derudover er der en database, der opsummerer de fysiologiske egenskaber af 152 methanogene isolater, tilgængelig1 (Jabłoński et al., 2015). For nylig afslørede genome binning usædvanlige mcrA-sekvenser i den nye klasse Methanofastidiosa (Nobu et al., 2016) og de nye fylaer Bathyarchaeota (Evans et al., 2015) og Verstraetearchaeota (Vanwonterghem et al., 2016). Disse nye fund udvidede vores viden om mangfoldigheden af potentielle methanogener, men de gjorde det ikke tvivl om anvendeligheden af mcrA-genet som en molekylær markør for det store flertal af methanogene samfund.
Til dato er methanogene samfund blevet påvist i vådområder, sedimenter, permafrostområder, rismarker, rådnetanke, geotermiske kilder og hydrotermiske slamslukningssteder (Conrad, 2007; Thauer et al., 2008; Wagner og Liebner, 2009). Det blev konstateret, at den methanogene samfundsstruktur var forbundet med miljøets pH-værdi, temperatur, saltholdighed, grundvandsniveau og vegetationsdynamik på forskellige rumlige og tidsmæssige skalaer (Megonigal et al., 2005; Milferstedt et al., 2010; Frank-Fahle et al., 2014; McCalley et al., 2014; Cui et al., 2015; Liebner et al., 2015). F.eks. er acetoklastisk metanogenese generelt hæmmet af lav pH, da det reducerer acetatdissociationen (Megonigal et al., 2005; Kotsyurbenko et al., 2007). Vegetationen kan levere labilt organisk kulstof af høj kvalitet som brændstof til methanogenerne i form af rodekssudater eller detritus, således at planteekssudater generelt begunstiger acetoklastiske methanogener primært i højmoser (Bridgham et al., 2013). Sulfat fra havvand hæmmer metanproduktionen i vådområder med tidevand, og saltholdighed er derfor blevet anvendt som en generel prædiktor for metanemissioner (Holm et al., 2016). En undersøgelse af tibetanske søers sedimenter viste, at stigende saltholdighed hæmmer hydrogenotrofiske methanogener, men øger acetoklastisk methanogenese (Liu et al., 2016). Disse undersøgelser indikerede miljømæssige drivkræfter for methanogene samfund, men har fokuseret på enkelte habitater eller begrænsede rumlige skalaer.
For at forstå methanogeners tilpasning til forskellige miljøændringer er det imidlertid nødvendigt med en systematisk og global udforskning af sammenhængene mellem mikrobielle samfundssammensætning og miljøforhold (Lozupone og Knight, 2007). På nuværende tidspunkt er der kun få undersøgelser, der omhandler spredning og habitatfiltrering af methanogene samfund (Auguet et al., 2010; Barreto et al., 2014). Vi opstiller den hypotese, at methanogene sammensætninger hovedsageligt påvirkes af habitatfiltrering, og at den er drevet af globale miljøkontroller. I betragtning af at metanemissionshastighederne er meget forskellige mellem naturlige økosystemer, kan den eksplicitte integration af sammensætningen, diversiteten og biogeografien af methanogene assemblages i disse økosystemer være grundlæggende for at bestemme metanproduktionens reaktion på nuværende og fremtidige klimaændringer. Denne metastudie er udført for at udfylde hullerne i forbindelse med methanogen biogeografi, diversitet og dens miljømæssige kontrol ved hjælp af offentligt tilgængelige mcrA-sekvensdata og litteratur suppleret med fysiologiske data fra methanogene isolater.
Materialer og metoder
Dataindsamling
Vi hentede mcrA-sekvenser, der er tilgængelige i GenBank (januar 2016)2. For hvert hit blev den originale artikel kontrolleret, og de tilsvarende mcrA-sekvenser blev analyseret af et brugerdefineret Perl-script. Da vi fokuserede på naturlige miljøer, blev methanogene mcrA-sekvenser indhentet fra naturlige habitater og klassificeret som jord, sø-, flodmundings-, hav- og hydrotermiske sedimenter og muddervulkaner. Fem biblioteker fra næste generations sekventering (NGS) blev inkluderet ud over sekvenser fra klonbiblioteker. Sekvenserne blev downloadet uden at tage hensyn til den relative hyppighed i det oprindelige datasæt. Da sekvenser af klonbiblioteker hovedsagelig dækker de rigelige fylotyper, mens NGS kan indfange en meget dybere diversitet, gik vi på kompromis med at anvende NGS-dataene, men afbød en potentiel fejl på grund af sekventeringsmetodernes forskellige opløsning. Derfor valgte vi kun de repræsentative sekvenser af rigelige OTU’er med en relativ hyppighed på mere end 1 %. Vi afviste desuden de NGS-sekvenser, som ikke bestod oversættelseskontrollen fra nukleotid- til proteinsekvenser, eller som havde en lav kvalitet (sekvenser < 250 bps). Endelig konstruerede vi et datasæt, der indeholdt 4466 unikke mcrA-sekvenser fra 94 globalt fordelte steder (figur 1 og supplerende tabeller S1, S2). Desuden trak vi ikke mcrA-sekvenser af potentielle arkæale methanotrofer fra datasættet, hvilket uundgåeligt blev opdaget i den genomiske undersøgelse (Conrad, 2007). Denne del ligger uden for fokus i denne undersøgelse.
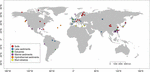
FIGUR 1. Placering af de lokaliteter, der er anvendt i denne undersøgelse. Lokaliteterne blev inddelt i seks forskellige kategorier i henhold til deres levesteder og blev mærket med forskellige farver. En oversigt over de seks habitater findes i de supplerende tabeller S1, S2.
De geografiske koordinater og miljømæssige indstillinger, herunder pH, saltholdighed, højde, gennemsnitlig årlig lufttemperatur (MAAT) og gennemsnitlig årlig nedbør (MAP), blev for hvert forskningssted, der er taget i betragtning i denne undersøgelse, uddraget fra de tilsvarende publikationer, forudsat at dataene er tilgængelige (se supplerende tabel S1). For at tage hensyn til manglende miljøparametre på tværs af flere undersøgelser definerede vi kvalitativt nogle miljøvariabler i henhold til stedbeskrivelserne i den relevante litteratur og konverterede derefter disse kategoridata til semimetriske numeriske værdier, f.eks. definerede vi marine sedimenter, hydrotermiske sedimenter, vulkansk mudder og sodasø-sedimentprøver som “salte”, jord- og ferskvandssø-sedimentprøver som “ikke-saltede” og mangrove- og flodmundingsprøver som “blandede” prøver.
Raw Sequence Processing
Sekvensbehandlingen blev gennemført med Mothur-softwareplatformen (Schloss et al, 2009). Sekvenser fra forskellige biblioteker blev sammenlagt før behandlingen. Sekvenser med en længde på mindre end 350 bp eller mere end 8 tvetydige baser blev kasseret. Efterfølgende blev disse sekvenser justeret i forhold til en forudjusteret delmængde af mcrA-sekvenser, som blev hentet fra FunGene-databasen på http://fungene.cme.msu.edu/ (Fish et al., 2013). Chimeriske sekvenser blev identificeret med Mothur-softwaren ved hjælp af uchime-metoden (Edgar et al., 2011) med selve datasættet som reference. Derefter blev de gyldige mcrA-gen-nukleotidsekvenser anvendt til at beregne ukorrigerede parvise afstande mellem alignede DNA-sekvenser og yderligere tildelt i operationelle taksonomiske enheder (OTU) ved en cutoff på 84 %, der svarer til 97 % for 16S rRNA-genet (Yang et al., 2014). Hyppigheden af hver mcrA OTU blev kun opgjort som tilstedeværelse eller fravær. Vi øgede nøjagtigheden af den taksonomiske klassificering af OTU’erne ved at tage hensyn til både nukleotid- og aminosyresekvenser. På DNA-niveau blev den taksonomiske identitet tildelt af Mothur-platformen i henhold til en referencedatabase (Yang et al., 2014). På proteinniveau blev de tilpassede proteinsekvenser brugt til at konstruere et træ i ARB, og derefter blev den taksonomiske tildeling baseret på den tilsvarende database. Hvis tilknytningen af en OTU var inkonsistent, blasted vi manuelt både nukleotid- og proteinsekvensen i NCBI og bestemte den endelige taksonomiske identitet ved at tage hensyn til forespørgselsdækningen (>95%), identiteten (>84%) og e-værdien (<1E-5). For proteinsekvenser henviste cutoff på slægtsniveau til tærsklen på 83,5% (Hunger et al., 2011).
Økologisk og statistisk analyse
Den statistiske analyse blev udført af forskellige R-pakker. Principal coordinates analysis (PCoA)-ordinationer blev genereret på grundlag af Jaccard-afstandsmatricer, der blev konstrueret ved hjælp af vegan-pakken v2.2.0 (Oksanen et al., 2015). Permutational MANOVA (multivariat variansanalyse) blev udført for at vurdere kilden til variation i Jaccard-matrixen (McArdle og Anderson, 2001) i vegan med 104 permutationer. Jaccard-afstandsmålingerne er baseret på artens tilstedeværelse/fravær, hvilket er mere egnet til vores datasæt, da de fleste undersøgelser kun leverede de repræsentative sekvenser, mens oplysningerne om abundans mangler. De taksonomiske forekomsthyppigheder på tværs af habitater blev visualiseret ved hjælp af bubble plots med ggplot2-pakken (v1.0.0.0) (Wickham, 2009). Hierarkisk klyngeanalyse af de ikke-saltede jord- og søsedimentsamfund blev udført ved hjælp af R-funktionen ‘hclust’ (R Core Team, 2014). De opnåede samfundsklynger blev beskrevet i henhold til de oprindelige prøvers pH- og temperaturregime, fordi vi ved hjælp af PCoA på forhånd havde identificeret begge parametre pH og temperatur som havende indflydelse på sammensætningen af methanogene samfund i ikke-saltede jordarter og søsedimenter. Tilknytningen af hver methanogen slægt til hver af disse klynger blev bestemt ved hjælp af korrelationsbaseret indikatorartsanalyse (Dufrene og Legendre, 1997). Indikatorarter er her defineret som arter, der både forekommer hyppigt i en bestemt type levested (specificitet) og overvejende findes i denne type levested (trofasthed). I denne undersøgelse blev indikatortaxaerne, svarende til indikatorartsbegrebet, for ikke-saltjord og søsedimenter udvalgt i henhold til en indikatorværdi (IndVal-værdi) af R-pakken labdsv (Roberts, 2016), hvis sandsynligheden for at opnå en indikatorværdi så høj som den observerede i løbet af de angivne iterationer er mindre end 0,05. Chao2-indeksene blev beregnet for hver prøve ved hjælp af pakken vegan. Wilcoxon-ranksummetesten af Chao2-indeks mellem habitater blev udført ved hjælp af R-funktionen “wilcox.test” (R Core Team, 2014). For at konsolidere virkningen af habitatfiltrering på methanogene archaea-isolater blev de fysiologiske og biokemiske karakteristika for beskrevne methanogene kulturer hentet fra3 ‘Methanogenic archaea database’ (Jabłoński et al., 2015). Blandt dem blev isolaterne med oplysninger om optimalt NaCl-behov filtreret, kategoriseret og plottet i henhold til deres isolationskilde.
For at undersøge indflydelsen af spredningsbegrænsning på methanogene samfundets struktur blev der udført en lineær regressionsanalyse baseret på en geografisk afstandsmatrix og en Jaccard-afstandsmatrix for samfund ved hjælp af R-funktionen “lm” (R Core Team, 2014). Vi udførte Mantel- og partielle Mantel-tests for at evaluere virkningerne af spredningsbegrænsning i henhold til de to matricer igen ved hjælp af vegan-pakken i R (Oksanen et al., 2015). Endvidere blev multivariat rumlig analyse (spatial PCA) anvendt på 16 europæiske jord- og søsedimentprøver baseret på Morans I-indeks for at undersøge den rumlige struktur af methanogener ved hjælp af funktionen “multispati” i R ade4-pakken (Dray og Dufour, 2007). Desuden blev Ward’s Minimum variance clustering, som var baseret på Jaccard-distancematrixen, implementeret på disse 16 prøver ved hjælp af R-funktionen “hclust” (R Core Team, 2014), og vi projicerede yderligere clusteringresultaterne på et geografisk kort. Den europæiske shapefile til kortlægning på statsniveau er tilgængelig på GSHHG-databasen (v2.3.6)4 . Kortet blev genereret ved hjælp af QGIS v2.18.25.
Resultater
Biogeografi af methanogene arkæa i naturlige miljøer
Der blev hentet mcrA-gensekvenser fra 94 globalt distribuerede naturlige miljøer. Placeringen og økosystemtypen af hvert af disse 94 steder er afbildet i figur 1. Incidensfrekvenserne (tilstedeværelse/fravær) af methanogene slægtslinjer blev slået sammen i henhold til økosystemtype og illustreret i figur 2. Kort fortalt er Methanoregula det hyppigst observerede taxon i jordbunden sammen med Methanobacterium, Methanosaeta, Methanocella, Methanomassiliicoccus og Methanosarcina. I sedimenter fra flodmundinger blev der hyppigt fundet sekvenser fra Methanosaeta, Methanobacterium, Methanoregula og Methanoculleus. I søsedimenter var det desuden hovedsagelig Methanoregula og Methanosaeta, der blev fundet. I marine sedimenter er Methanoculleus og Methanosaeta de mest almindelige lineager, efterfulgt af Methanolinea.
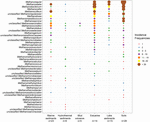
FIGUR 2. Bobleplot, der viser hyppigheden af forekomst af methanogene lineager i forskellige naturlige miljøer. Rangordenen langs den lodrette akse svarer til de faldende samlede incidensfrekvenser for linearerne. Taxonomien er vist for slægtsniveauet. Hvis det ikke var muligt at henføre til slægtsniveauet, blev det næste højere taksonomiske niveau anvendt. Antallet af prøver (n) er angivet for hvert levested.
Selv om mange taxa blev påvist i forskellige miljøer, viser nogle stadig miljøpræferencer. Methanoregula, det taxon, der hyppigt forekommer i ikke-marine miljøer og overgangsmiljøer (jord, søsedimenter og flodmundinger), er fraværende i marine habitater (marine sedimenter, hydrotermiske sedimenter og muddervulkaner). Methanobacterium og Methanocella, som er fremherskende i ikke-marine miljøer og overgangsmiljøer, findes sjældent i marine levesteder. I modsætning hertil er Methanococcoides, som er en fremherskende slægt i marine sedimenter, hydrotermiske sedimenter og muddervulkaner, næsten ikke observeret i jord og søsedimenter. Desuden er Methanogenium og Methanolacinia kun observeret i flodmundinger og marine sedimenter, mens Methanospirillum og Methanosphaerula kun er fundet i terrestriske miljøer. Desuden findes nogle specifikke taxa udelukkende i hydrotermiske sedimenter, herunder Methanocaldococcus, Methanothermococcus, Methanopyrus, Methanotorris og Methanococcus. Selv om nogle slægter som Methanosaeta er til stede i de fleste miljøer, kan ingen slægt betragtes som allestedsnærværende.
Den største rigdom af slægter fandtes i sedimenter fra flodmundinger, som også rummer mere jævne forekomsthyppigheder af forskellige slægter. I modsætning hertil viser muddervulkaner og hydrotermiske økosystemer en relativt lav methanogen diversitet. Jord og søsedimenter, der ligner estuarer, er karakteriseret ved forskellige methanogene assemblages.
Alpha-diversitet af methanogene samfund i naturlige miljøer
Rigdommen af methanogene archaea i henhold til Chao2-indekset varierede i høj grad mellem økosystemtyperne (figur 3). Med henblik på direkte sammenligning af alfa-diversiteterne og for at opnå en rimelig afvejning mellem prøverne blev der foretaget en subsampling til 15 sekvenser for hvert sted. Chao2-indekset viser, at sedimenter fra flodmundinger omfatter den største artsrigdom af methanogener blandt de seks økosystemtyper (Supplerende tabel S3), hvilket understreger resultaterne af bobleplottet (figur 2). Jord- og søsedimenter, der viser en lavere artsrigdom end østersøprøver, har signifikant højere Chao2-indeks end marine sedimenter og hydrotermiske sedimenter og uden signifikante forskelle mellem marine sedimenter og hydrotermiske sedimenter (Supplerende tabel S3).
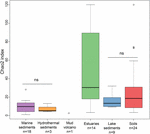
FIGUR 3. Boksdiagram af Chao2-indeks for de forskellige økosystemtyper. Plottet er baseret på undersamplede datasæt, der indeholder 15 sekvenser for hvert sted, for at gøre sammenligningen på alfa-diversitetsmål mere robust. Antallet af prøver i hvert habitat er angivet som “n” under habitatbetegnelsen. “ns” angiver ingen statistisk signifikans i Wilcoxon-testen. Det statistiske resultat af alfa-diversitet på OTU-niveau er angivet i Supplerende tabel S3.
Globale kontroller af methanogene samfund i naturlige miljøer
De 94 globalt distribuerede methanogene samfund blev grupperet i et ordinationsplot ved at anvende PCoA baseret på Jaccard-afstandsmatrixen. Ifølge PCoA-analysen forklarer den første og anden akse tilsammen 16,3 % af den samlede varians. Variationerne mellem prøverne kan således i vid udstrækning forklares af saltholdigheden (figur 4). Da der i nogle tilfælde ikke forelå nogen indledende data om saltkoncentrationerne, har vi kvalitativt klassificeret disse prøver som saltvandsprøver, blandede (mellemliggende) prøver og ikke-saltvandsprøver som beskrevet ovenfor. De saltholdige og ikke-saltholdige prøver adskilles effektivt langs den første akse. De blandede prøver er samlet set grupperet mellem saltvandsprøverne og de ikke-saltvandsprøverne. Den permutationelle MANOVA baseret på Jaccard-afstandsmatrixen tyder også på, at saltholdighed er den primære abiotiske faktor, der styrer fordelingen af globale methanogene samfund (R2 = 0,099, P < 0,001) (tabel 1).
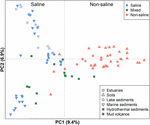
FIGUR 4. Principal coordinates analysis (PCoA) ordination baseret på Jaccard-afstandsmatrixen af methanogene mcrA-gen-sekvensbiblioteker med sammenligning af 94 prøver. PCoA’en er farvet efter saltholdighed: De røde symboler angiver ikke-saltvandsmiljøer, de blå angiver saltvandsmiljøer, og de grønne angiver mellemliggende miljøer. Forskellige symboler viser forskellige miljøer. Procentdelen af den variation, der forklares af de plottede hovedkoordinater, er angivet på akserne.

TABEL 1. Permutationel MANOVA-analyse på Jaccard-afstandsmatrixen af alle prøverne fra de seks habitater for at teste sammenhængen mellem samfundets varians og forskellige miljøvariabler.
Dertil kommer, at vi kontrollerede for en potentiel sammenhæng mellem isoleringskilden for methanogene renkulturer og den optimale koncentration af NaCl til vækst. Den optimale NaCl-koncentration for de methanogene renkulturer viste et fald fra marine til flodmundinger, søsedimenter og jordøkosystemer (Supplerende figur S2). Nogle få isolater med en outlier stammer fra sodasø-sedimenter eller hypersaline jorde.
Miljøkontrol og methanogene indikatortaxa i ikke-saltede jorde og søsedimenter
På globalt plan er methanogene samfund fra ikke-saltede jorde og søsedimenter tæt sammenbragt (figur 4), så vi analyserede yderligere miljøkontrollerne af methanogene samfund fra disse to habitater, som udgør 33 undersøgelsessteder i alt. En fællesskabsbaseret klyngeanalyse for disse to typer habitater afslørede fire klynger baseret på Jaccard-afstandsmatrixen (Supplerende figur S3). Permutationel MANOVA tyder på, at både pH (R2 = 0,099, P < 0,001) og temperatur (R2 = 0,069, P < 0,001) påvirker den methanogene β-diversitet i ikke-saltede jordtyper og søsedimenter (tabel 2). I overensstemmelse hermed tildelte vi de fire klynger til pH og MAAT på det oprindelige prøvetagningssted og opnåede stort set konsistente undergrupper til samfundsklyngen (figur 5C,D). Kombinationen af miljøkarakteristika og disse fire samfundsklynger gør det muligt for os at definere disse fire undergrupper som neutral og kold gruppe 1, sur og kold gruppe 2, sur og moderat gruppe 3 og neutral og varm gruppe 4 (figur 5). Yderligere PCoA-ordination baseret på Jaccard-dissimilaritetsmatrixen viser, at langs PC1-aksen er de fleste prøver fra gruppe 2 og gruppe 3 fra sure jorde og søsedimenter, mens gruppe 1 og gruppe 4 hovedsagelig var fra neutrale miljøer (Figur 5A). Desuden adskilte prøverne fra moderate lokaliteter (gruppe 3) sig fra prøverne fra varme og kolde lokaliteter langs PC2, mens prøverne fra de varme miljøer (gruppe 4) adskilte sig fra de andre prøver langs PC3 (figur 5B). Dermed forklarer de tre første akser i PCoA-ordinationen 38,8 % af den samlede variation.

TABEL 2. Permutationel MANOVA baseret på en Jaccard-afstandsmatrix af prøver af ikke-saltjord og søsedimentprøver for at teste sammenhængen mellem samfundets varians og forskellige miljøvariabler.
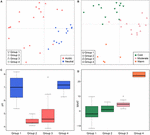
FIGUR 5. Plot for hovedkoordinatanalyse baseret på en Jaccard-afstandsmatrix for 33 ikke-saltede jorde og søsedimenter. Underplot (A) viser PC1 og PC2, og symbolerne er farvet efter pH, og underplot (B) viser PC2 og PC3, og symbolerne er farvet efter temperatur. De tre første komponenter forklarer 17,1, 11,7 og 10 % af variansen. Boxplotterne viser pH (C) og MAAT (gennemsnitlig årlig lufttemperatur) (D) for fire identificerede undergrupper. Farven på boksen i figur C svarer til pH-kategorien i (A). Farverne i boksdiagrammerne viser statistisk signifikans baseret på en parvis Wilcoxon-test (P < 0,05), hvor prøverne med samme farve ikke adskiller sig signifikant fra hinanden. På samme måde følger farven i (D) temperaturgrupperingen i (B). Undergrupperne henviser til den hierarkiske klyngeanalyse af samfundsligheder.
Vi undersøgte forekomsten af methanogene lineager i hver undergruppe baseret på incidensfrekvenser (figur 6). Methanoregula er fremherskende i alle typer ikke-saltvandshabitater. Ud over Methanoregula viser den neutrale og kolde undergruppe (gruppe 1) en høj forekomst af Methanosaeta, Methanobacterium og Methanosarcina. Den sure og kolde gruppe 2 er repræsenteret af Methanobacterium, Methanocella og Methanosarcina, mens Methanosaeta er fraværende her. Methanocella og Methanosaeta er almindelige i den sure og moderate gruppe 3. I den neutrale og varme gruppe 4 er Methanolinea og Methanosaeta vigtige medlemmer. Denne gruppe er den eneste gruppe, hvor Methanoculleus blev identificeret.

FIGUR 6. Incidensfrekvenser af methanogene slægtslinjer inden for de fire definerede grupper af methanogene samfund i jord og søsedimenter. Den lodrette akse er ordnet i alfabetisk rækkefølge. Søjlelængden svarer til de gennemsnitlige incidensfrekvenser for hver lineage inden for den tilsvarende gruppe. Fejlstængerne repræsenterer standardafvigelsen for et givet taxon i de forskellige prøver i den pågældende gruppe. Asteriskerne viser de specialiserede taxa med P-værdi < 0,05. Taxonomien er vist for slægtsniveauet. Hvis en tildeling til slægtsniveauet ikke var mulig, blev det næsthøjeste taxonomiske niveau, der kunne tildeles, anvendt. Beskrivelsen for de fire grupper fremgår af figur 5.
For alle fire grupper er de taxa, der har en høj forekomst, Methanoregula, Methanobacterium, Methanosarcina, Methanosaeta, Methanomassiliicoccus og Methanocella. De specialiserede taxa, som er betydeligt mere repræsenteret på de fleste lokaliteter inden for en given gruppe, blev fundet ved hjælp af indikatorartsanalyser som beskrevet ovenfor. I alt viste seks ud af de 31 taxa en signifikant indikatorværdi (P < 0,05) (markeret med asterisk i figur 6). Gruppe 1 (neutral og kold) viste det største antal specialister med slægtslinjer af Methanosaeta, Methanolobus og Methanomethylovorans. Methanobacterium fungerede som specialiseret taxon i den kolde og sure gruppe 2, mens Methanolinea blev identificeret som specialist i gruppe 4 (neutral og varm), men blev næsten ikke observeret i de andre grupper. Desuden er Methanoregula i høj grad repræsenteret i den sure og moderate gruppe 3.
Dispersal Limitation
En lineær regressionsanalyse (R2= 0,05, P < 0,001) viste en svag korrelation mellem geografisk afstand og methanogene samfundsstruktur på det globale datasæt. Samtidig viste en Mantel-test, at miljøvariablerne har en højere korrelation med samfundsstrukturen end de geografiske afstande (se tabel 3). Denne tendens bekræftes også af en delvis Mantel-test, der kontrollerer for autokorrelationseffekter. Plottet af den geografiske afstand i forhold til Jaccard-samfundets lighed viser ingen klar lineær tendens, men mønstre, der hovedsagelig skyldes den globale fordeling af prøveudtagningsstederne (se supplerende figur S4).

TABEL 3. Mantel- og partielle Mantel-testanalyser til bestemmelse af indflydelsen af miljøvariabler og geografisk afstand på den mikrobielle fordeling for det globale datasæt og en delprøve af 16 europæiske prøver.
For yderligere at analysere indflydelsen af spredning begrænsede vi vores analyse til Europa, hvor der blev udtaget de tætteste og mest jævne prøver. Mantel-tests og partielle Mantel-tests på denne delmængde reproducerede den tendens, at samfundsdataene er højere korreleret med miljøvariablerne end med geografiske afstande (se tabel 3). Den delvise Mantel-test, der kontrollerede for miljøvariabler, kunne ikke påvise nogen statistisk signifikant korrelation mellem det mikrobielle samfund og den geografiske afstand. En rumlig PCA-analyse af disse 16 europæiske lokaliteter viser, at der er en rumlig struktur i det methanogene samfund (23,7 % af den samlede varians blev forklaret af denne struktur), hvilket svarer til en positiv rumlig autokorrelation mellem lokaliteterne som angivet ved Morans I-indekset (Morans I = 0,4018). Kun den første egenværdi var stabil og svarede til en adskillelse af prøverne mellem Centraleuropa og de baltiske lande (Supplerende figur S5). Det lille datasæt vanskeliggør imidlertid en robust tildeling af denne observerede rumlige struktur til geografiske eller miljømæssige variabler eller til begge dele. Vi foretog derfor en klyngeanalyse af de methanogene samfund som beskrevet ovenfor og afslørede tre grupper (figur 7A), som vi projicerede på et geografisk kort (figur 7B). Klyngeopdelingen reproducerede ikke adskillelsen af den rumlige PCA langs Østersøen. Nogle lokaliteter, som geografisk set ligger meget tæt på hinanden, udviser således forskellige methanogene samfundsstrukturer og samles i forskellige klynger. På den anden side viser nogle geografisk meget fjerntliggende lokaliteter meget ens samfundssammensætninger og grupperer sig sammen (figur 7B). Den regionale spredning af grupperne i Europa synes ikke at svare til eller være begrænset af en geografisk struktur.
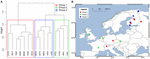
FIGUR 7. Hierarkisk klyngeanalyse af Jaccard-afstandsmatrixen blandt 16 europæiske jord- og søsedimentprøver. Ward-grupperingsmetoden blev anvendt til analysen. Lokaliteterne blev inddelt i tre grupper farvet af forskellige rektangler i klyngedendrogrammet (A) og blev projiceret på et europæisk kort (B).
Diskussion
Identificering og anvendelse af biogeografiske begreber på mikrobielle samfund er af stor interesse inden for mikrobiel økologi. Mikrobiel biogeografi menes at være styret af det evolutionære og økologiske samspil mellem fire hovedprocesser: habitatfiltrering, spredning, drift og mutation (Hanson et al., 2012). Selv om indflydelsen af drift og mutation ligger uden for denne undersøgelses fortolkningsmuligheder, viser vi, at der findes globale mønstre af methanogene samfund i naturlige miljøer. Denne undersøgelse viser et globalt biogeografisk mønster af metanogene samfund, som er mere forbundet med habitatfiltrering end med geografisk spredning. Methanogene samfund fra sedimenter fra sodasøer er f.eks. tæt forbundet med geografisk fjerntliggende marine prøver (figur 4), og meget ens methanogene samfund forekommer i europæiske jord- og søsedimenter på trods af stor afstand mellem hinanden (figur 7). Samlet set afslørede vores forsøg på at afdække potentielle spredningsbegrænsninger en svag indflydelse af geografisk placering på metanogene samfundsstrukturer, hvilket står i modsætning til en klar indflydelse af miljøforhold. Det er ikke muligt at udelukke en endelig udelukkelse af rumlige virkninger på de mikrobielle samfund med de tilgængelige datapunkter. Prøveudtagningsstederne fokuserer på bestemte habitater og/eller områder, mens det samlede antal prøver er lavt. Hvis der er en effekt, antager vi, at de rumlige effekter kun forekommer på regionalt eller lokalt plan. Der er rapporteret om lokale spredningsbegrænsninger for hypertermofile archaea forårsaget af geografiske barrierer (Whitaker et al., 2003), ammoniak-oxiderende bakterier på lokale saltmarskområder, men ikke på regionale og kontinentale skalaer (Martiny et al., 2011), mikroorganismer i dybhavssedimenter sammen med miljømæssige forhold (Schauer et al., 2009), og for bakterier i et stort sæt heterogene snemiljøer, som hovedsagelig skyldes tilgængeligheden af allochtonisk kulstof (Lutz et al., 2016). En mindre indflydelse af spredningsbegrænsning på methanogener i naturlige miljøer betyder, at methanogener kan fordele sig tilfældigt over rummet, udvælges med succes af det lokale miljø, hvis deres fysiologiske krav er opfyldt, og kan etablere stabile samfund (Martiny et al., 2006; von Mering et al., 2007).
Begrebet habitatfiltrering indebærer, at arter med lignende økologiske krav bør forekomme sammen oftere end forventet ved en tilfældighed (Weiher og Keddy, 2001; Cornwell et al., 2006; Ulrich et al., 2010). Vores resultat viser, at der er store forskelle i sammensætningen af methanogene samfund mellem marine økosystemer og økosystemer i vådområder og søer, mens flodmundinger placerer sig midt imellem. Biogeografiske mønstre baseret på et sæt genundersøgelser på miljøprøver blev også rapporteret for generelle bakterier (Nemergut et al., 2011), mikrobiomet for metan-slip (Ruff et al., 2015), ammoniak-oxiderende arkæa (Cao et al., 2013), marine pelagiske og bentiske bakterier (Zinger et al., 2011) og kvælstofcyklende mikroorganismer (Church et al., 2008). Habitatfiltrering blev specifikt rapporteret for ukulturerede archaea (Auguet et al., 2010), hele bakteriesamfund i forskellige miljøer (von Mering et al., 2007; Chaffron et al., 2010) eller i sydamerikanske tørveområder på regional skala (Oloo et al., 2016), såvel som specifikke bakteriegrupper såsom metanoxiderende (Knief, 2015) og kvælstoffikserende bakterier (Nelson et al., 2016).
Vores resultater viser, at på global skala regulerer saltholdigheden i væsentlig grad sammensætningen af methanogene samfund og bestemmer store forskelle mellem marine og terrestriske methanogene sammensætninger. Også methanogener fra sodasø-sedimenter klynger sig sammen med dem fra marine sedimenter (figur 4), hvilket fremhæver den globale indflydelse af saltholdighed. Dette resultat er i overensstemmelse med andre undersøgelser baseret på 16S rRNA-genet, der afslører, at saltholdighed er en primær faktor, der former de globale mønstre for de samlede bakterie- og arkæalsamfund (Lozupone og Knight, 2007; Auguet et al., 2010; Caporaso et al., 2011; Cao et al., 2013). En lav indflydelse af geografisk adskillelse, men en stærk indvirkning af saltholdighed på de generelle mikrobielle samfund blev også observeret i tidligere undersøgelser (Logares et al., 2013; Yang et al., 2016). Følgelig bestemmer saltholdigheden i høj grad, hvilke lineager der kan overleve. I forskellige habitater var metanproduktionsaktiviteten negativt korreleret med saltholdigheden (Bartlett et al., 1987; Potter et al., 2009; Poffenbarger et al., 2011). Hæmningen af metanproduktionen på grund af saltholdighed foreslås dermed at falde sammen med en reduceret metanogen populationsstørrelse (Pattnaik et al., 2000). Effekten af saltholdighed på hydrogenotrofisk, acetotrofisk og methylotrofisk methanogenese afhænger dermed af saltholdighedsniveauet og er forskellig for de forskellige veje af methanogenese (Liu et al., 2016). På nuværende tidspunkt er der ingen klar mekanisme til at forklare saltholdighedens indvirkning på samfundsstrukturen, men flere hypoteser kan tjene som mulige forklaringer. Fysiologisk set påvirker saltholdighed cellernes eksterne og interne osmolaritet. De ikke-saltholdige methanogene celler har udviklet fysiologiske tilpasninger til at modvirke det interne turgortryk, mens de salttilpassede celler har mistet denne egenskab (Zinder, 1993). Desuden kan stigende saltholdighed få methanogenerne til at syntetisere eller optage en øget andel af kompatible solutater til en betydelig energi- og dermed metabolisk omkostning (McGenity, 2010). Egenskaben salttolerance manifesterer sig endda i den optimale NaCl-koncentration for vækst af methanogene renkulturer, da vi fandt, at isolaterne fra marine sedimenter og hydrotermiske sedimenter har en betydeligt højere optimal NaCl-koncentration end isolaterne fra jord (Supplerende figur S2).
I de ikke-saltede terrestriske økosystemer, specifikt i jord og søsedimenter, styres det methanogene samfundssammensætning af kombinationen af temperatur og pH. I overensstemmelse hermed kan methanogenerne i disse miljøer klassificeres i fire grupper (figur 6). I modsætning til marine økosystemer viser de ikke-saltede terrestriske økosystemer en stor naturlig variabilitet i både pH og temperatur. Temperaturen kan påvirke ikke blot den metanogene vej, men også de metanogene populationer selv (Conrad, 2007; Rooney-Varga et al., 2007). Metanproduktionen kan øges betydeligt, hvis temperaturen stiger som følge af de temperaturfølsomme trin under fermentering og acetogenese (Megonigal et al., 2005; Kotsyurbenko et al., 2007). Desuden kan lav pH-værdi i væsentlig grad begrænse tilgængeligheden af acetat ved at forhindre acetat i at dissocieres og dermed påvirke acetoklastisk metanogenese negativt (Fukuzaki et al., 1990; Bridgham et al., 2013). Dette kunne være en mulig årsag til, at Methanosaeta var fraværende i gruppe2, mens Methanosarcina kan skifte mellem forskellige kilder og ikke blev påvirket væsentligt. Desuden kan pH regulere effektiviteten af metanproduktionen og de metanogene veje fra ombrotrofe til minerotrofe tørveområder gennem direkte hæmning af begge metanogeneseveje og indirekte gennem dens virkninger på fermenteringen (Ye et al., 2012). Derfor kan både temperatur og pH direkte eller indirekte regulere metaboliske trin i forbindelse med methanogenese og den opstrøms fermentering, som leverer substrat til methanogenerne.
Methanoregula er allestedsnærværende og meget rigelig i alle fire grupper af terrestriske habitater (figur 6), men stort set fraværende i det marine system og kan således vise sig at være en proxy for ferskvandsindflydelse i det marine område. Dens globale relevans blev for nylig rapporteret andetsteds (Yang et al., 2017). På trods af sin allestedsnærværende udbredelse i jord og søsedimenter forekommer Methanoregula som en indikatorlinje i sure habitater med moderate temperaturer. Desuden synes (1) Methanolinea at være særligt tilpasset de varme, neutrale terrestriske miljøer, (2) Methanobacterium til de kolde, sure miljøer og (3) Methanosaeta til pH-neutrale miljøer, hvilket er i overensstemmelse med andre undersøgelser (Rosenberg et al., 2014) og understreger robustheden af vores tilgang. Generelt vil de geokemiske forhold omkring methanogene samfund føre til nichedifferentiering. Da nichesortering har tendens til at efterlade de adaptive specialister (Langenheder og Székely, 2011), genererede den progressive langsigtede miljøvalg på lang sigt en række nicher, der blev udfyldt af en række endemiske habitatspecialister, som kan være mindre repræsenteret eller fraværende i andre forskellige miljøforhold. Samfundet er også formet af biotiske faktorer, såsom økologiske interaktioner, dynamik, konkurrence og symbiose. På trods af disse biologiske faktorer fandt von Mering et al. (2007), at habitatpræferencer ofte er bemærkelsesværdigt stabile over tid, og den karakteristiske taksonomiske sammensætning af miljøsamfund kan til gengæld være en vigtig indikator for deres økologi og funktion.
I overensstemmelse med habitatpræferencen hos methanogene arkæer ser det ud til, at nært beslægtede methanogene stammer ofte blev isoleret fra sammenlignelige miljøer. For eksempel synes Methanoregulaceae at være ret forskelligartet i naturlige miljøer (Yang et al., 2017), og neutrale stammer kan indtil videre modstå dyrkning. De aktuelt beskrevne stammer af Methanoregula er begge opnået fra svagt acidofile miljøer, mens de to repræsentanter for Methanolinea er fra relativt varme habitater såsom henholdsvis rådnetøjsslam og rismarkjord (Rosenberg et al., 2014). Selv om Methanobacterium-stammer blev isoleret fra forskellige miljøer, udviser ca. halvdelen af de eksisterende isolater af denne slægt pH-optima lidt mindre end 7. Det betyder, at indikatorlinjerne, som blev identificeret på grundlag af miljøsekvenserne, kunne afspejle differentieringerne af fysiologi og kilder i de eksisterende methanogene kultivarer. For eksempel kan habitatets saltholdighed, som en generel egenskab ved habitatet, gradvist udsætte organismer for stærk miljømæssig udvælgelse og filtrere sammensætningen af et nyt sæt arter, som er bedst egnet til den omgivende saltholdighed (Logares et al., 2013).
Biodiversitetsbevaring og -forvaltning er en primær udfordring i vores nuværende samfund. Her viser vi, at methanogene archaea i naturlige miljøer er mest forskelligartede i flodmundingssedimenter. Estuarier er overgangszoner mellem marine og terrestriske økosystemer. Dette giver mulighed for to vigtige processer, der kan bidrage til artsrigdommen. For det første blandes mikrober fra havet og landjorden i flodmundingerne og omfatter i sidste ende en samlet høj diversitet (McLusky og Elliott, 2004). Der er f.eks. blevet observeret en høj diversitet af bakterier, arkæer, svampe og endda specifikke bakterier, der udfører unikke funktioner (Cunliffe et al., 2008; Mosier og Francis, 2008; Crump et al., 2012). Et andet aspekt er det høje næringsstofniveau som følge af de terrestriske, tidevandsbetingede tilførsler, som organerne i flodmundingen kan ernære sig af (McLusky og Elliott, 2004; Statham, 2012). I denne sammenhæng er flodmundingsmiljøerne af betydning for genvinding af generisk nyhed for methanogener. Indtil videre har virkningerne af artsdiversitet på økosystemprocesser tiltrukket en betydelig forskningsindsats. Sammenhængen mellem biodiversitet og økosystemfunktion er stadig under debat og er stadig uklar for mikrobielle samfund (Loreau et al., 2001; Tilman et al., 2014). Selv om jord og søsedimenter er primære kilder til metan og også habitater med høj metanogen diversitet, foreslår vi, at artsrigdom ikke er en passende proxy for metanproduktionspotentialet og økosystemets metanemissioner; den synes snarere at afspejle miljøets heterogenitet og historie. En rangordning af miljøerne efter deres artsrigdom er ikke nødvendigvis ensbetydende med potentialet for metanemissionsrater, som er højest fra jord og søer og forholdsvis mindre fra flodmundinger (figur 3 og supplerende figur S1).
Endeligt kan manglen på miljøoplysninger i de offentlige databaser have hindret en fuldstændig fortolkning af de miljømæssige drivkræfter, der er observeret her. Selv om der er stigende sekventeringsdata om methanogener i litteraturen og offentlige databaser, er de relaterede abiotiske variabler, der leveres, ofte inkonsekvente og sparsomme. Den begrænsede mængde af konsistente oplysninger om miljøvariabler begrænser anvendelsen af statistiske multivariate analyser. Når man husker på, at de abiotiske faktorer i denne undersøgelse kun kan forklare en begrænset del af varianserne i samfundet, tyder det på, at der mangler andre forklarende variabler. Af særlig betydning kunne være koncentrationerne og tilgængeligheden af methanogene substrater som f.eks. acetat, hydrogen og methylaminer. Ikke desto mindre vil abiotiske parametre måske aldrig være tilstrækkelige til fuldt ud at forklare, hvad der strukturerer methanogene sammensætninger, simpelthen fordi levestederne har forskellige historier og kun kan undersøges lokalt. Blandt de tilgængelige mcrA-data er de store områder i det russiske og canadiske subarktiske og arktiske område også dårligt repræsenteret. En bedre geografisk dækning og en jævn fordeling af mcrA-gen-datasæt ville forbedre vurderingen af metanogene samfund på globalt plan.
Author Contributions
SL og XW har designet undersøgelsen. XW og SY indsamlede og analyserede dataene. XW, SY og FH udførte den statistiske analyse. XW, MW og SY foretog den fylogenetiske korrektion. XW, SY, FH, MW, DW og SL fortolkede resultaterne og skrev artiklen. Alle forfattere bidrog til diskussionerne og gennemgik manuskriptet.
Interessekonflikter
Forfatterne erklærer, at forskningen blev udført uden kommercielle eller økonomiske relationer, der kunne opfattes som en potentiel interessekonflikt.
Akkreditering
Den økonomiske støtte til XW (Grant No. 201408620031 til XW), der blev ydet af China Scholarship Council (CSC), er taknemmelig. Denne undersøgelse blev støttet af Helmholtz Gemeinschaft (HGF) ved at finansiere Helmholtz Young Investigators Group of SL (VH-NG-919).
Supplementært materiale
Det supplerende materiale til denne artikel kan findes online på: https://www.frontiersin.org/article/10.3389/fmicb.2017.01339/full#supplementary-material
Fodnoter
- ^ http://metanogen.biotech.uni.wroc.pl/
- ^ http://www.ncbi.nlm.nih.gov/
- ^ http://metanogen.biotech.uni.wroc.pl/
- ^ https://www.ngdc.noaa.gov/mgg/shorelines/gshhs.html
- ^ http://qgis.osgeo.org
Auguet, J. C., Barberan, A., og Casamayor, E. O. (2010). Globale økologiske mønstre i ukulturerede Archaea. ISME J. 4, 182-190. doi: 10.1038/ismej.2009.109
PubMed Abstract | CrossRef Full Text | Google Scholar
Bange, H. W., Bartell, U. H., U. H., Rapsomanikis, S. og Andreae, M. O. (1994). Metan i Østersøen og Nordsøen og en revurdering af de marine emissioner af metan. Glob. Biogeochem. Cycles 8, 465-480. doi: 10.1029/94GB02181
CrossRef Full Text | Google Scholar
Barreto, D. P., Conrad, R., Klose, M., Claus, P., og Enrich-Prast, A. (2014). Distance-decay og taxa-arealrelationer for bakterier, archaea og methanogene archaea i et tropisk søsediment. PLoS ONE 9:e110128. doi: 10.1371/journal.pone.0110128
PubMed Abstract | CrossRef Full Text | Google Scholar
Bartlett, K. B., Bartlett, D. S., D. S., Harriss, R. C., og Sebacher, D. I. (1987). Metanemissioner langs en saltholdighedsgradient i en saltmarsk. Biogeochemistry 4, 183-202. doi: 10.1007/BF02187365
CrossRef Full Text | Google Scholar
Borrel, G., O’Toole, P. W., Harris, H. M. B., Peyret, P., Brugere, J. F., and Gribaldo, S. (2013). Fylogenomiske data understøtter en syvende orden af methylotrofiske methanogener og giver indsigt i udviklingen af methanogenese. Genome Biol. Evol. 5, 1769-1780. doi: 10.1093/Gbe/Evt128
PubMed Abstract | CrossRef Full Text | Google Scholar
Bousquet, P., Ciais, P., Miller, J., Dlugokencky, E., Hauglustaine, D., Prigent, C., et al. (2006). Bidrag fra antropogene og naturlige kilder til atmosfæriske metanvariationer. Nature 443, 439-443. doi: 10.1038/nature05132
PubMed Abstract | CrossRef Full Text | Google Scholar
Bridgham, S. D., Cadillo-Quiroz, H., Keller, J. K., and Zhuang, Q. (2013). Metanemissioner fra vådområder: biogeokemiske, mikrobielle og modelleringsperspektiver fra lokale til globale skalaer. Glob. Chang. Biol. 19, 1325-1346. doi: 10.1111/gcb.12131
PubMed Abstract | CrossRef Full Text | Google Scholar
Cao, H., Auguet, J.-C., and Gu, J.-D., og Gu, J.-D. (2013). Globalt økologisk mønster af ammoniak-oxiderende arkæaer. PLoS ONE 8:e52853. doi: 10.1371/journal.pone.0052853
PubMed Abstract | CrossRef Full Text | Google Scholar
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Lozupone, C. A., Turnbaugh, P. J., et al. (2011). Globale mønstre af 16S rRNA-diversitet med en dybde på millioner af sekvenser pr. prøve. Proc. Natl. Acad. Sci. U.S.A. 108, 4516-4522. doi: 10.1073/pnas.1000080107
PubMed Abstract | CrossRef Full Text | Google Scholar
Chaffron, S., Rehrauer, H., Pernthaler, J., og von Mering, C. (2010). Et globalt netværk af sameksisterende mikrober ud fra miljø- og helgenomsekvensdata. Genome Res. 20, 947-959. doi: 10.1101/gr.104521.109
PubMed Abstract | CrossRef Full Text | Google Scholar
Church, M. J., Bjorkman, K. M., Karl, D. M., D. M., Saito, M. A., og Zehr, J. P. (2008). Regional fordeling af kvælstoffikserende bakterier i Stillehavet. Limnol. Oceanogr. 53, 63-77. doi: 10.4319/lo.2008.53.1.0063
CrossRef Full Text | Google Scholar
Conrad, R. (2007). “Microbial ecology of methanogens and methanotrophs,” in Advances in Agronomy, ed. L. S. Donald (Cambridge, MA: Academic Press), 1-63.
Google Scholar
Cornwell, W. K., Schwilk, D. W., og Ackerly, D. D. (2006). En egenskabsbaseret test for habitatfiltrering: konveks skrogvolumen. Ecology 87, 1465-1471. doi: 10.1890/0012-9658(2006)872.0.CO;2
PubMed Abstract | CrossRef Full Text | Google Scholar
Crump, B. C., Amaral-Zettler, L. A., og Kling, G. W. (2012). Mikrobiel diversitet i arktiske ferskvand er struktureret af inokulering af mikrober fra jordbunden. ISME J. 6, 1629-1639. doi: 10.1038/ismej.2012.9
PubMed Abstract | CrossRef Full Text | Google Scholar
Cui, M., Ma, A., Qi, H., Zhuang, X., Zhuang, G., og Zhao, G. (2015). Varmere temperatur fremskynder metanemissioner fra Zoige-vådområdet på den tibetanske højslette uden at ændre sammensætningen af methanogene samfund. Sci. Rep. 5:11616. doi: 10.1038/srep11616
PubMed Abstract | CrossRef Full Text | Google Scholar
Cunliffe, M., Schafer, H., Harrison, E., Cleave, S., Upstill-Goddard, R., og Murrell, J. C. (2008). Fylogenetisk og funktionel genanalyse af bakterie- og archaealsamfundene i forbindelse med overflademikrolaget i et flodmundingsområde. ISME J. 2, 776-789. doi: 10.1038/ismej.2008.28
PubMed Abstract | CrossRef Full Text | Google Scholar
Dray, S., and Dufour, A. B. (2007). Pakken ade4: implementering af dualitetsdiagrammet for økologer. J. Stat. Softw. 22, 1-20. doi: 10.18637/jss.v022.i04
CrossRef Full Text | Google Scholar
Dufrene, M., og Legendre, P. (1997). Artsammensætninger og indikatorarter: behovet for en fleksibel asymmetrisk tilgang. Ecol. Monogr. 67, 345-366. doi: 10.2307/2963459
CrossRef Full Text | Google Scholar
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C., og Knight, R. (2011). UCHIME forbedrer følsomheden og hastigheden af chimera-detektion. Bioinformatics 27, 2194-2200. doi: 10.1093/bioinformatics/btr381
PubMed Abstract | CrossRef Full Text | Google Scholar
Evans, P. N., Parks, D. H., Chadwick, G. L., Robbins, S. J., Orphan, V. J., Golding, S. D., et al. (2015). Metanmetabolisme i archaeal phylum Bathyarchaeota afsløret af genom-centreret metagenomik. Science 350, 434-438. doi: 10.1126/science.aac7745
PubMed Abstract | CrossRef Full Text | Google Scholar
Fish, J. A., Chai, B., Wang, Q., Sun, Y., Brown, C. T., Tiedje, J. M., et al. (2013). FunGene: den funktionelle gen-pipeline og -opbevaring. Front. Microbiol. 4:291. doi: 10.3389/fmicb.2013.00291
PubMed Abstract | CrossRef Full Text | Google Scholar
Frank-Fahle, B. A., Yergeau, E., Greer, C. W., Lantuit, H., og Wagner, D. (2014). Mikrobielt funktionelt potentiale og samfundssammensætning i permafrostpåvirkede jorde i det nordvestlige canadiske arktiske område. PLoS ONE 9:e84761. doi: 10.1371/journal.pone.0084761.g001
PubMed Abstract | CrossRef Full Text | Google Scholar
Fukuzaki, S., Nishio, N., og Nagai, S. (1990). Kinetik af den methanogene fermentering af acetat. Appl. Environ. Microbiol. 56, 3158-3163.
Google Scholar
Hanson, C. A., Fuhrman, J. A., Horner-Devine, M. C., og Martiny, J. B. (2012). Ud over biogeografiske mønstre: processer, der former det mikrobielle landskab. Nat. Rev. Microbiol. 10, 497-506. doi: 10.1038/Nrmicro2795
PubMed Abstract | CrossRef Full Text | Google Scholar
Holm, G. O., Perez, B. C., McWhorter, D. E., Krauss, K. W., Johnson, D. J., Raynie, R. C., et al. (2016). Metanstrømme på økosystemniveau fra ferskvands- og brakvandssumpområder i Mississippi River Delta: implikationer for kulstofprojekter i kystnære vådområder. Wetlands 36, 401-413. doi: 10.1007/s13157-016-0746-7
CrossRef Full Text | Google Scholar
Hunger, S., Schmidt, O., Hilgarth, M., Horn, M. A., Kolb, S., Conrad, R., et al. (2011). Konkurrerende formater- og kuldioxid-brugende prokaryoter i en anoxisk metanemitterende mosejord. Appl. Environ. Microbiol. 77, 3773-3785. doi: 10.1128/AEM.00282-11
PubMed Abstract | CrossRef Full Text | Google Scholar
IPCC (2007). “Climate Change 2007: the Physical Science Basis”, i Proceedings of the Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, eds D. Qin, S. Solomon, M. Manning, Z. Chen, M. Marquis, K. B. Averyt, et al. (Cambridge: Cambridge University Press).
Google Scholar
Jabłoński, S., Rodowicz, P., og Łukaszewicz, M. (2015). Methanogen archaea-database indeholdende fysiologiske og biokemiske karakteristika. Int. J. Syst. Evol. Microbiol. 65, 1360-1368. doi: 10.1099/ijs.0.000065
PubMed Abstract | CrossRef Full Text | Google Scholar
Kirschke, S., Bousquet, P., Ciais, P., Saunois, M., Canadell, J. G., Dlugokencky, E. J., et al. (2013). Tre årtier med globale metankilder og -sænkninger. Nat. Geosci. 6, 813-823. doi: 10.1038/Ngeo1955
CrossRef Full Text | Google Scholar
Knief, C. (2015). Diversitet og habitatpræferencer hos dyrkede og uopdrættede aerobe methanotrofe bakterier vurderet på baggrund af pmoA som molekylær markør. Front. Microbiol. 6:1346. doi: 10.3389/fmicicb.2015.01346
PubMed Abstract | CrossRef Full Text | Google Scholar
Kotsyurbenko, O., Friedrich, M., Simankova, M., Nozhevnikova, A., Golyshin, P., Timmis, K., et al. (2007). Skift fra acetoklastisk til H2-afhængig methanogenese i en vestsibirisk tørvemose ved lave pH-værdier og isolering af en acidofil Methanobacterium-stamme. Appl. Environ. Microbiol. 73, 2344-2348. doi: 10.1128/AEM.02413-06
PubMed Abstract | CrossRef Full Text | Google Scholar
Langenheder, S., og Székely, A. J. (2011). Sortering af arter og neutrale processer er begge vigtige under den indledende samling af bakteriesamfund. ISME J. 5, 1086-1094. doi: 10.1038/ismej.2010.207
PubMed Abstract | CrossRef Full Text | Google Scholar
Liebner, S., Ganzert, L., Kiss, A., Yang, S., Wagner, D., og Svenning, M. M. (2015). Skift i methanogene samfundssammensætning og metanfluxer langs nedbrydningen af diskontinuerlig permafrost. Front. Microbiol. 6:356. doi: 10.3389/fmicb.2015.00356
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, Y. Q., Priscu, J. C., Xiong, J. B., Conrad, R., Vick-Majors, T., Chu, H. Y., et al. (2016). Salinitet driver archaeal distributionsmønstre i højhøjde søsedimenter på den tibetanske højslette. FEMS Microbiol. Ecol. 92:fiw033. doi: 10.1093/femsec/fiw033
PubMed Abstract | CrossRef Full Text | Google Scholar
Logares, R., Lindstrom, E. S., Langenheder, S., Logue, J. B., Paterson, H., Laybourn-Parry, J., et al. (2013). Biogeografi af bakteriesamfund, der er udsat for progressive langsigtede miljøændringer. ISME J. 7, 937-948. doi: 10.1038/ismej.2012.168
PubMed Abstract | CrossRef Full Text | Google Scholar
Loreau, M., Naeem, S., Inchausti, P., Bengtsson, J., Grime, J. P., Hector, A., et al. (2001). Biodiversitet og økosystemers funktion: nuværende viden og fremtidige udfordringer. Science 294, 804-808. doi: 10.1126/science.1064088
PubMed Abstract | CrossRef Full Text | Google Scholar
Lozupone, C. A., and Knight, R. (2007). Globale mønstre i bakteriel diversitet. Proc. Natl. Acad. Sci. U.S.A. 104, 11436-11440. doi: 10.1073/pnas.0611525104
PubMed Abstract | CrossRef Full Text | Google Scholar
Luton, P. E., Wayne, J. M., Sharp, R. J., og Riley, P. W. (2002). mcrA-genet som et alternativ til 16S rRNA i den fylogenetiske analyse af methanogenpopulationer i deponeringsanlæg. Microbiology 148, 3521-3530. doi: 10.1099/00221287-148-11-3521
PubMed Abstract | CrossRef Full Text | Google Scholar
Lutz, S., Anesio, A. M., Raiswell, R., Edwards, A., Newton, R. J., Gill, F., et al. (2016). Biogeografien af røde snemikrobiomer og deres rolle i smeltende arktiske gletsjere. Nat. Commun. 7:11968. doi: 10.1038/ncomms11968
PubMed Abstract | CrossRef Full Text | Google Scholar
Martiny, J. B. H., Bohannan, B. J. M. M., Brown, J. H., Colwell, R. K., Fuhrman, J. A., Green, J. L., et al. (2006). Mikrobiologisk biogeografi: at sætte mikroorganismer på kortet. Nat. Rev. Microbiol. 4, 102-112. doi: 10.1038/Nrmicro1341
PubMed Abstract | CrossRef Full Text | Google Scholar
Martiny, J. B. H., Eisen, J. A., Penn, K., Allison, S. D., og Horner-Devine, M. C. (2011). Drivers of bacterial beta-diversity depend on spatial scale. Proc. Natl. Acad. Sci. U.S.A. 108, 7850-7854. doi: 10.1073/pnas.1016308108
PubMed Abstract | CrossRef Full Text | Google Scholar
McArdle, B. H., og Anderson, M. J. (2001). Tilpasning af multivariate modeller til samfundsdata: en kommentar til afstandsbaseret redundansanalyse. Ecology 82, 290-297. doi: 10.1890/0012-9658(2001)0822.0.CO;2
CrossRef Full Text | Google Scholar
McCalley, C. K., Woodcroft, B. J., Hodgkins, S. B., Wehr, R. A., Kim, E.-H., Mondav, R., et al. (2014). Metan-dynamik reguleret af mikrobielle samfunds reaktion på permafrost optøning. Nature 514, 478-481. doi: 10.1038/nature13798
PubMed Abstract | CrossRef Full Text | Google Scholar
McGenity, T. J. (2010). “Methanogens and Methanogenesis in Hypersaline Environments,” in Handbook of Hydrocarbon and Lipid Microbiology, ed. K. N. Timmis (Berlin: Springer), 665-680. doi: 10.1007/978-3-540-77587-4_53
CrossRef Full Text | Google Scholar
McLusky, D. S., og Elliott, M. (2004). Det østatlige økosystem: Økologi, trusler og forvaltning. New York, NY: Oxford University Press. doi: 10.1093/acprof:oso/9780198525080.001.0001
CrossRef Full Text | Google Scholar
Megonigal, J. P., Mines, M. E., og Visscher, P. T. (2005). “Anaerobic metabolism: linkages to trace gases and aerobic processes,” in Biogeochemistry, ed. W. H. Schlesinger (Oxford: Elsevier), 350-362.
Google Scholar
Milferstedt, K., Youngblut, N. D., og Whitaker, R. J. (2010). Rumlig struktur og persistens af methanogenpopulationer i humiske mosesøer. ISME J. 4, 764-776. doi: 10.1038/Ismej.2010.7
PubMed Abstract | CrossRef Full Text | Google Scholar
Mosier, A. C., og Francis, C. A. (2008). Relativ hyppighed og diversitet af ammoniak-oxiderende archaea og bakterier i San Francisco-bugtens flodmunding. Environ. Microbiol. 10, 3002-3016. doi: 10.1111/j.1462-2920.2008.01764.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Nazaries, L., Murrell, J. C., Millard, P., Baggs, L., and Singh, B. K. (2013). Metan, mikrober og modeller: grundlæggende forståelse af jordens metancyklus med henblik på fremtidige forudsigelser. Environ. Microbiol. 15, 2395-2417. doi: 10.1111/1462-2920.12149
PubMed Abstract | CrossRef Full Text | Google Scholar
Nelson, M. B., Martiny, A. C., og Martiny, J. B. H. (2016). Global biogeografi af mikrobielle kvælstofcyklus-egenskaber i jord. Proc. Natl. Acad. Sci. U.S.A. 113, 8033-8040. doi: 10.1073/pnas.1601070113
PubMed Abstract | CrossRef Full Text | Google Scholar
Nemergut, D. R., Costello, E. K., Hamady, M., Lozupone, C., Jiang, L., Schmidt, S. K., et al. (2011). Globale mønstre i biogeografien af bakterietaxa. Environ. Microbiol. 13, 135-144. doi: 10.1111/j.1462-2920.2010.02315.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Nobu, M. K., Narihiro, T., Kuroda, K., Mei, R., og Liu, W. T. (2016). Jagten på den flygtige Euryarchaeota-klasse WSA2: genomer afslører en unik fastidiøs methylreducerende methanogen. ISME J. 10, 2478-2487. doi: 10.1038/ismej.2016.33
PubMed Abstract | CrossRef Full Text | Google Scholar
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., P. R., O’Hara, R. B., et al. (2015). vegan: Community Ecology Package. R Package Version 2.2.0. Tilgængelig på: https://CRAN.R-project.org/package=vegan
Google Scholar
Oloo, F., Valverde, A., Quiroga, M. V., Vikram, S., Cowan, D., and Mataloni, G. (2016). Habitatheterogenitet og konnektivitet former mikrobielle samfund i sydamerikanske tørveområder. Sci. Rep. 6:25712. doi: 10.1038/srep25712
PubMed Abstract | CrossRef Full Text | Google Scholar
Pattnaik, P., Mishra, S. R., Bharati, K., Mohanty, S. R., Sethunathan, N. og Adhya, T. K. (2000). Indflydelse af saltholdighed på metanogenese og den tilhørende mikroflora i tropiske risjorde. Microbiol. Res. 155, 215-220. doi: 10.1016/S0944-5013(00)80035-X
PubMed Abstract | CrossRef Full Text | Google Scholar
Poffenbarger, H. J., Needelman, B. A., og Megonigal, J. P. (2011). Salinitetens indflydelse på metanemissioner fra tidevandssumpområder. Wetlands 31, 831-842. doi: 10.1007/s13157-011-0197-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Potter, E. G., Bebout, B. M., og Kelley, C. A. (2009). Isotopsammensætningen af metan og afledte metanogene substrater langs en saltholdighedsgradient i et hypersalt mikrobielt mattsystem. Astrobiology 9, 383-390. doi: 10.1089/ast.2008.0260
PubMed Abstract | CrossRef Full Text | Google Scholar
R Core Team (2014). R: A Language and Environment for Statistical Computing (version 3.0.3). Wien: R Foundation for Statistical Computing.
Google Scholar
Roberts, D. (2016). labdsv: Ordination og multivariat analyse for økologi. R Package Version 1.8-0. Tilgængelig på: https://CRAN.R-project.org/package=labdsv
Google Scholar
Rooney-Varga, J. N., Giewat, M. W., Duddleston, K. N., Chanton, J. P., and Hines, M. E. (2007). Sammenhænge mellem archaeal-samfundsstruktur, vegetationstype og metanogene veje i tørveområder i Alaska. FEMS Microbiol. Ecol. 60, 240-251. doi: 10.1111/j.1574-6941.2007.00278.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Rosenberg, E., DeLong, E. F., Lory, S., Stackebrandt, E., og Thompson, F. (2014). The Prokaryotes: Andre større bakterielinjer og archaea. Berlin: Springer.
Google Scholar
Ruff, S. E., Biddle, J. F., Teske, A. P., Knittel, K., Boetius, A., og Ramette, A. (2015). Global spredning og lokal diversificering af mikrobiomet i metanudsivninger. Proc. Natl. Acad. Sci. U.S.A. 112, 4015-4020. doi: 10.1073/pnas.1421865112
PubMed Abstract | CrossRef Full Text | Google Scholar
Schauer, R., Bienhold, C., Ramette, A., og Harder, J. (2009). Bakteriediversitet og biogeografi i dybhavsoverfladesedimenter i det sydlige Atlanterhav. ISME J. 4, 159-170. doi: 10.1038/ismej.2009.106
PubMed Abstract | CrossRef Full Text | Google Scholar
Schloss, P. D., Westcott, S. L., Ryabin, T., Hall, J. R., Hartmann, M., Hollister, E. B., et al. (2009). Introduktion af mothur: open source, platform-uafhængig, fællesskabsstøttet software til beskrivelse og sammenligning af mikrobielle samfund. Appl. Environ. Microbiol. 75, 7537-7541. doi: 10.1128/Aem.01541-09
PubMed Abstract | CrossRef Full Text | Google Scholar
Statham, P. J. (2012). Næringsstoffer i flodmundinger – en oversigt og de potentielle virkninger af klimaændringer. Sci. Total Environ. 434, 213-227. doi: 10.1016/j.scitotenv.2011.09.088
PubMed Abstract | CrossRef Full Text | Google Scholar
Thauer, R. K., Kaster, A.-K., Seedorf, H., Buckel, W., og Hedderich, R. (2008). Methanogene arkæaer: økologisk relevante forskelle i energibesparelse. Nat. Rev. Microbiol. 6, 579-591. doi: 10.1038/Nrmicro1931
PubMed Abstract | CrossRef Full Text | Google Scholar
Tilman, D., Isbell, F., and Cowles, J. M. (2014). Biodiversitet og økosystemers funktion. Annu. Rev. Ecol. Evol. Syst. 45, 471-493. doi: 10.1146/annurev-ecolsys-120213-091917
CrossRef Full Text | Google Scholar
Ulrich, W., Hajdamowicz, I., Zalewski, M., Stañska, M., Ciurzycki, W., og Tykarski, P. (2010). Artssortering eller habitatfiltrering: en casestudie af edderkoppesamfund på øer i søer. Ecol. Res. 25, 375-381. doi: 10.1007/s11284-009-0661-y
CrossRef Full Text | Google Scholar
Vanwonterghem, I., Evans, P. N., Parks, D. H., Jensen, P. D., Woodcroft, B. J., Hugenholtz, P., et al. (2016). Methylotrofisk methanogenese opdaget i den arkæale phylum Verstraetearchaeota. Nat. Microbiol. 1:16170. doi: 10.1038/nmicrobiol.2016.170
PubMed Abstract | CrossRef Full Text | Google Scholar
von Mering, C., Hugenholtz, P., Raes, J., Tringe, S. G., Doerks, T., Jensen, L. J., et al. (2007). Kvantitativ fylogenetisk vurdering af mikrobielle samfund i forskellige miljøer. Science 315, 1126-1130. doi: 10.1126/science.1133420
PubMed Abstract | CrossRef Full Text | Google Scholar
Wagner, D., and Liebner, S. (2009). “Global warming and carbon dynamics in permafrost soils: methane production and oxidation,” in Permafrost Soils, ed. R. Margesin (Berlin: Springer), 219-236.
Google Scholar
Weiher, E., og Keddy, P. (2001). Økologiske forsamlingsregler: Perspektiver, fremskridt, tilbagetrækninger. Cambridge: Cambridge University Press.
Google Scholar
Whitaker, R. J., Grogan, D. W., og Taylor, J. W. (2003). Geografiske barrierer isolerer endemiske populationer af hypertermofile archaea. Science 301, 976-978. doi: 10.1126/science.1086909
PubMed Abstract | CrossRef Full Text | Google Scholar
Wickham, H. (2009). ggplot2: Elegant Graphics for Data Analysis (Elegant grafik til dataanalyse). New York, NY: Springer-Verlag. doi: 10.1007/978-0-387-98141-3
CrossRef Full Text | Google Scholar
Yang, J., Ma, L. A., Jiang, H., Wu, G., og Dong, H. (2016). Salinitet former den mikrobielle diversitet og samfundsstruktur i overfladesedimenter i Qinghai-Tibetan-søerne. Sci. Rep. 6:25078. doi: 10.1038/srep25078
PubMed Abstract | CrossRef Full Text | Google Scholar
Yang, S., Liebner, S., Alawi, M., Ebenhöh, O., og Wagner, D. (2014). Taxonomisk database og cut-off-værdi til behandling af mcrA-genet 454-pyrosequencingdata ved MOTHUR. J. Microbiol. Methods 103, 3-5. doi: 10.1016/j.mimet.2014.05.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Yang, S., Liebner, S., Winkel, M., Alawi, M., Horn, F., Dörfer, C., et al. (2017). Dybdegående analyse af methanogene kernesamfund fra permafrostpåvirkede vådområder i høj højde. Soil Biol. Biochem. 111, 66-77. doi: 10.1016/j.soilbio.2017.03.007
CrossRef Full Text | Google Scholar
Ye, R. Z., Jin, Q. S., Bohannan, B., Keller, J. K., McAllister, S. A., og Bridgham, S. D. (2012). pH-kontrol over anaerob kulstofmineralisering, effektiviteten af metanproduktion og methanogene veje i tørvemoser på tværs af en ombrotrofisk-minerotrofisk gradient. Soil Biol. Biochem. 54, 36-47. doi: 10.1016/j.soilbio.2012.05.015
CrossRef Full Text | Google Scholar
Zinder, S. H. (1993). “Fysiologisk økologi af methanogener”, i Methanogenese: Ecology, Physiology, Biochemistry & Genetics, ed. J. G. Ferry (Boston, MA: Springer), 128-206.
Google Scholar
Zinger, L., Amaral-Zettler, L. A., Fuhrman, J. A., Horner-Devine, M. C., Horner-Devine, M. C., Huse, S. M., Welch, D. B. M., et al. (2011). Globale mønstre af bakteriel beta-diversitet i økosystemer på havbunden og i havvand. PLoS ONE 6:e24570. doi: 10.1371/journal.pone.0024570
PubMed Abstract | CrossRef Full Text | Google Scholar