Plasmamembraner
Plasmamembranen er en struktur i plantecellen, der danner en semipermeabel eller selektiv barriere mellem cellens indre og det ydre miljø; den fungerer også ved transport af molekyler ind i og ud af cellen.
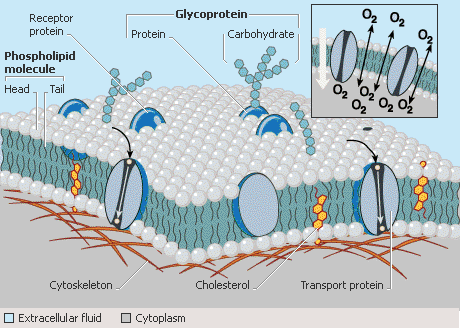
Ud over at danne den strukturelle barriere mellem cellens indre indhold og det ydre miljø indeholder plasmamembraner proteiner, der er involveret i transport af molekyler og andre stoffer ind og ud af cellen, og de indeholder proteiner og andre molekyler, der er vigtige for at modtage signaler fra omgivelserne og fra plantehormoner, der styrer vækst og deling.
Kulhydrater, der er forbundet med plasmamembranen, er markører for celletypen. Hos planter er plasmamembranen stedet for cellulosesyntese.


Lipidmolekyler giver struktur til plasmamembranen, som beskrives af den flydende mosaikmodel som et dynamisk hav af lipider, hvori andre molekyler flyder.
Phospholipider er den hyppigst forekommende lipid i plasmamembraner, og de er organiseret i et flydende fosfolipid-dobbeltlag, hvori steroler, proteiner og andre molekyler er spredt ud. Fosfolipider er amfipatiske molekyler, der indeholder vandelskende (hydrofile) områder og vandskyende (hydrofobiske) områder.
Hvert fosfolipid består af en glycerolryggekæde med tre kulstofatomer; to af kulstofferne er knyttet til langkædede fedtsyremolekyler, og det tredje kulstof er knyttet til en fosfatholdig gruppe. Da fedtsyrerne er upolære og hydrofobiske, har de en tendens til at aggregere og udelukke vand.
Denne aggregering gør det muligt for fosfolipiderne at danne en dobbeltlagsstruktur, der har fedtsyrerne i begge lag i midten og de ladede, fosfatholdige grupper mod ydersiden.
Denne tolagestruktur gør det muligt for den ene overflade af plasmamembranens tolag at interagere med det vandige ydre miljø, mens den anden interagerer med det vandige indre cellemiljø.
Steroler findes også i plasmamembranerne i planteceller. Det vigtigste sterol, der findes i plantecellers plasmamembraner, er stigmasterol (i modsætning til kolesterol, som findes i dyrecellers plasmamembraner). Steroler, der findes i planteceller, er økonomisk vigtige som udgangsmateriale for steroidbaserede lægemidler som f.eks. p-piller.
Membranproteiner og kulhydrater
Nogle membranproteiner spænder over hele længden af fosfolipiddobbeltlaget og kaldes transmembranproteiner. Transmembranproteiner kaldes undertiden for integrale membranproteiner og har forskellige strukturer og funktioner.
De kan kun passere gennem lipiddobbeltlaget én gang, eller de kan være “multiple pass” transmembranproteiner, der væver sig ind og ud af membranen mange gange.
Den del af et transmembranprotein, der passerer gennem membranens indre, består ofte af aminosyrer, der har upolære sidekæder (R-grupper), og er kendt som transmembrandomænet.
Den del af trans-membranproteinet, der befinder sig på membranens ydre overflade og interagerer med det vandige miljø, indeholder ofte ladede, eller polære, aminosyrer i sin sekvens.
Membranproteiner er ofte vigtige for at modtage signaler fra det ydre miljø som membranreceptorer. F.eks. interagerer protein- eller peptidhormoner med transmembranproteinreceptorer på plasmamembranen. Membranproteiner er også involveret i at modtage signaler som f.eks. lysfotoner.
Membranproteiner danner porer, der tillader ioner (ladede partikler) at passere gennem det indre af membranen. Membranproteiner, der kaldes carriers, er vigtige for at bringe næringsmolekyler som f.eks. simple sukkerstoffer ind i cellen.
Det er ikke alle proteiner i membranen, der er transmembranproteiner. Nogle er kun løst forbundet med membranen, er knyttet til andre proteiner eller er forankret i membranen med en lipidhale. Disse proteiner,som ikke spænder over begge sider af membranen, kaldes ofte perifere membranproteiner.
Ud over proteiner indeholder plasmamembranen kulhydratmolekyler. Kulhydratmolekyler er normalt knyttet til membranproteiner eller til lipidmolekyler i bilaget. Kulhydrater giver vigtig information om celletype og identitet.
Transport på tværs af membraner
Transport af molekyler ind i og ud af cellerne er en vigtig funktion for plasmamembranen. Hydrofobiske molekyler, som f.eks. ilt, og små, uladede molekyler, som f.eks. kuldioxid, krydser membranen ved simpel diffusion.
Disse molekyler bruger den potentielle energi fra en kemisk gradient til at drive deres bevægelse fra et område med højere koncentration på den ene side af membranen til et område med lavere koncentration på den anden side.
Diffusionen fungerer bedst, når denne koncentrationsgradient er stejl. I celler, der ikke har evnen til at udføre fotosyntese, bliver ilten f.eks. brugt næsten lige så hurtigt, som den kommer ind i cellen.
Derved opretholdes en skarp gradient af iltmolekyler på tværs af membranen, således atmolekyler kontinuerligt strømmer fra området med større iltkoncentration uden for cellen til området med lavere koncentration inde i cellen.
Molekyler, der er polære, udelukkes fra det hydrofobiske område af bilaget. To faktorer påvirker transporten af disse typer molekyler: koncentrationsgradienten og den elektriske gradient. Lipiddobbeltlag adskiller forskelle i elektrisk ladning fra den ene side af membranen til den anden, hvilket fungerer som en slags biologisk kondensator.
Hvis indersiden af cellen er mere negativ end ydersiden af cellen, vil negativt ladede ioner skulle bevæge sig fra indersiden til ydersiden af cellen for at rejse med den elektriske gradient. Kombinationen af koncentrationsgradienten og den elektriske gradient kaldes den elektrokemiske gradient.
Transport af ladede eller polære molekyler kræver hjælp fra proteiner i membranen, kendt som transportere. Kanalproteiner danner porer i membranen og tillader små, ladede molekyler, som regel uorganiske ioner, at strømme over membranen fra den ene side til den anden.
Hvis ionens bevægelsesretning er nedad dens elektrokemiske gradient, kræver processen ikke ekstra energi og kaldes passiv transport.
Transportproteiner ændrer form for at deponere et lille molekyle, f.eks. et sukker, fra den ene side af en membran til den anden. Pumper er proteiner i membranen, der bruger energi fra adenosintrifosfat (ATP) eller lys til at transportere molekyler over membranen. Når der bruges energi i transporten, kaldes processen for aktiv transport.
Cellulosebiosyntese
I planter er plasmamembranen stedet for syntesen af cellulose, det mest rigelige biopolymer på jorden. Elektronmikroskopiske undersøgelser tyder på, at plantecellemembranen indeholder rosetstrukturer, der er komplekser af mange proteiner, og som er stedet for cellulosesyntesen.
Studier i bakterier, bomuldsplanter og ukrudtet Arabidoposis thaliana har gjort det muligt for forskerne at isolere det gen, der rent faktisk udfører de kemiske reaktioner, der binder glukosemolekyler sammen til den lange cellulosemikrofibrille-struktur.
Dette gen koder for et protein kaldet glycosyltransferase. Antistoffer mod den katalytiske eller aktive underenhed af glycosyltransferase mærker specifikt disse rosetstrukturer.
To af disse transferase-molekyler virker samtidig fra modsatte sider for at tilføje to glukoser ad gangen til den voksende mikrofibrille, hvilket forklarer rotationen af vekslende glukoser i cellulosemolekylerne.