De fleste af os lærte i skolen, at der er tre slags levende pattedyr – eutherier, pungdyr og monotreme – og at de mest indlysende forskelle mellem dem er, hvordan de formerer sig. De eutheriske eller “placentale” pattedyr, som f.eks. mennesker, udgør langt størstedelen af nutidens pattedyrs mangfoldighed. Eutherier har alle en chorioallantoisk placenta, et bemærkelsesværdigt organ, der dannes efter befrugtningen på det sted, hvor embryoet kommer i kontakt med slimhinden i moderens livmoder (Langer, 2008).
Marsupialer og monotreme dyr håndterer graviditet forskelligt (Abbot og Rokas, 2017; Renfree, 2010). Æglæggende monotreme, som f.eks. det andeskæggede næbdyr, har små “pugler”, der klækkes fra læderagtige skaller. Marsupialer – kænguruer, koalaer, bandicoots, opossum og så videre – har levende fødsler, men deres graviditeter er korte, og deres små unger er udviklingsmæssigt umodne og har tilsyneladende ikke noget behov for en moderkage. Efter fødslen fortsætter joeyerne med at udvikle sig uden for moderens krop, ofte i folder og poser på moderens mave.
I pungdyr er den mælk, som moderen giver efter fødslen, central for afkommets udvikling, og i modsætning til hos eutheriske pattedyr ændrer sammensætningen af denne mælk sig dramatisk, efterhånden som de unge joeyer vokser. I det væsentlige udfører pungdyrenes mælkekirtler mange af de funktioner, som den eutheriske placenta har (Renfree, 2010; Sharp et al., 2017; figur 1). Og for at sætte det, du måske har lært i biologiundervisningen, endnu mere på hovedet, har pungdyr alligevel en moderkage, men den udvikles sent i graviditeten og fra andre væv end hos eutherier. Forskellen mellem eutheriske pattedyr og pungdyr er således ikke tilstedeværelsen eller fraværet af en placenta, men snarere den relative vægt, der lægges på placenta og amning for at pleje afkommet gennem udviklingen.
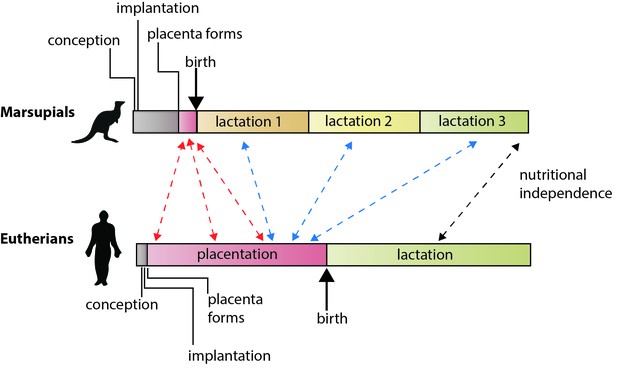
De forskellige reproduktionsstrategier hos eutheriske pattedyr og pungdyr
I eutheriske pattedyr er den energi, som moderen investerer i at opdrage ungerne før fødslen (via placenta) og efter fødslen (via amning), nogenlunde lige stor. Hos pungdyr er drægtigheden kort, placentaen dannes sent i drægtigheden, og amningen er langvarig. Guernsey et al. viser, at genetiske træk, der regulerer udviklingen via placenta hos eutherier, deles med den kortlivede placenta hos pungdyr (røde pile). De viser også, at nogle af de gener, der ligger til grund for placentafunktioner hos eutherier, udtrykkes under laktationen hos pungdyr (blå pile), herunder forskellige bevarede komponenter af selve laktationen (sort pil; Lefèvre et al., 2010). Bemærk: tidsskalaerne er ikke absolutte.
Nu, i eLife, rapporterer Julie Baker fra Stanford University School of Medicine, Marilyn Renfree fra University of Melbourne og medarbejdere – herunder Michael Guernsey fra Stanford som førsteforfatter, Edward Chuong fra University of Utah og Guillaume Cornelis (Stanford) – nye detaljer om de molekylære mekanismer, der ligger til grund for placentation og laktation hos eutherier og marsupials (Guernsey et al., 2017). Resultaterne blev opnået ved at bruge en modificeret version af en teknik kaldet RNA-seq til at måle, hvordan transkriptomet (det komplette sæt af RNA-transskriptioner i en celle eller et sæt af celler) varierede mellem forskellige celletyper i løbet af udviklingen (Rokas og Abbot, 2009).
Guernsey et al. sammenlignede ændringer i genekspression i to celletyper i placentaen hos en tammar wallaby, et lille australsk pungdyr, i løbet af udviklingen. De fandt, at genekspressionen var forskellig mellem de to væv, og at den desuden ændrede sig dynamisk over tid, svarende til det, der sker hos eutherier. Blandt transskriptionerne fandt de desuden mange, der havde kritiske funktioner i eutheriske placentaer, herunder medlemmer af Ig7-signalvejene og GCM1, en transkriptionsfaktor, der er vigtig for dannelsen og udviklingen af placentaen. Og det var ikke blot generne, der var bevaret, men også genekspressionsmønstrene i wallabyens moderkage lignede dem, der blev set i musens moderkage i de tidlige stadier af graviditeten. Det er præcis, hvad vi ville forvente at se, hvis moderkagen varetager tidlige udviklingsfunktioner hos wallabyen, og senere funktioner leveres efter fødslen. Dette tyder på, at en væsentlig forskel mellem pungdyr og eutherier ikke ligger i placentaens tidlige funktioner, men snarere i, hvordan placentafunktionerne er blevet opdelt i løbet af udviklingen af eutheriernes graviditet.
Sluttelig karakteriserede Guernsey et al. mønstrene af genekspression i tammars og flere pattedyrs brystkirtler. Både mus og wallaby delte lignende mønstre af genekspression, hvilket understregede temaet om funktionel kompartmentalisering og bevarelse i begge grupper. Men det mest bemærkelsesværdige er, at de identificerede en række gener, der udtrykkes i tammars brystkirtler, som vides at være funktionelt vigtige i placentaen hos eutherier (figur 1). Disse gener omfattede gener, der er involveret i transport af næringsstoffer, og flere gener, der vides at være nødvendige for placentation hos eutherier (herunder GCM1). Denne bevarelse af genekspression argumenterer for, at placentaen hos pungdyr styrer den tidlige fosterudvikling og laktationen styrer den sene fosterudvikling ved hjælp af nogle af de samme gener og molekylære veje som den eutheriske placenta.
De, der studerer pungdyr, har længe argumenteret for, at vi er nødt til at rette vores lærebøger for at anerkende pungdyrs placentaer og deres særprægede komplekse laktation (Renfree, 1983). Guernsey et al. styrker sagen ved at påvise, at både eutherier og pungdyr udtrykker en bevaret værktøjskasse af gener, der kan være lokaliseret til forskellige væv og organer, men som tjener fælles formål i fosterudviklingen. Denne overraskende bevarelse understreger vigtigheden af at identificere de gener, der ligger til grund for funktionelle ændringer i løbet af evolutionen (Rausher og Delph, 2015).
Hensyneladende er det værd at bemærke, at pungdyr varierer enormt i reproduktive træk (Tyndale-Briscoe, 2005), og at karakterisering af flere arter på den måde, som Guernsey et al. har gjort for tammar wallaby, vil give en rigere forståelse af evolutionen og diversiteten af selve pungdyrs drægtighed. Der er dog behov for mere arbejde for at udvikle passende statistiske metoder til kvantificering af bevarelsen af transkriptomprofiler mellem arter. Og hvis man ser ud over pattedyr, findes der former for placentation i alt fra øgler til søheste og insekter, og foreløbige undersøgelser tyder på, at mange af de involverede gener eller træk deles (Ostrovsky et al., 2016; Whittington et al., 2015). Det vil være fascinerende at finde ud af, hvor dybt vi kan spore oprindelsen af graviditetsværktøjskassen.