A legtöbbünk azt tanulta az iskolában, hogy az élő emlősöknek három fajtája van – az eutriák, az erszényesek és az egypatások -, és hogy a legnyilvánvalóbb különbség közöttük az, hogy hogyan szaporodnak. Az eutriák vagy “méhlepényes” emlősök, mint például az ember, alkotják a mai emlősök sokféleségének túlnyomó többségét. Az eutheriák mindegyike rendelkezik chorioallantoikus placentával, egy figyelemre méltó szervvel, amely a fogamzás után alakul ki azon a helyen, ahol az embrió kapcsolatba kerül az anya méhének bélésével (Langer, 2008).
Az erszényesek és az egypatások másképp kezelik a terhességet (Abbot és Rokas, 2017; Renfree, 2010). A tojást tojó egypatásoknak, mint például a kacsacsőrű tintahalaknak, apró “puglijaik” vannak, amelyek bőrszerű héjakból kelnek ki. Az erszényesek – a kenguruk, koalák, bandicootok, oposszumok stb. – élve szülnek, de terhességük rövid, és apró kölykeik fejlődésileg éretlenek, és úgy tűnik, nincs szükségük méhlepényre. A születés után a kisállatok az anyjuk testén kívül fejlődnek tovább, gyakran az anyjuk hasán lévő ráncokban és tasakokban.
A erszényeseknél az anya által a születés után adott tej központi szerepet játszik az utódok fejlődésében, és az egeres emlősöktől eltérően ennek a tejnek az összetétele drámaian megváltozik, ahogy a kisállatok növekednek. Lényegében az erszényesek emlőmirigyei az egerek méhlepényének számos funkcióját látják el (Renfree, 2010; Sharp et al., 2017; 1. ábra). És hogy még inkább felborítsuk a biológiaórán tanultakat, az erszényeseknek mégiscsak van méhlepényük, de az a terhesség késői szakaszában és más szövetekből fejlődik ki, mint az eutriáknál. Az eutriás emlősök és az erszényesek közötti különbség tehát nem a méhlepény megléte vagy hiánya, hanem a placenta és a szoptatás relatív hangsúlya, amelyet az utódok fejlődésének ápolására fordítanak.
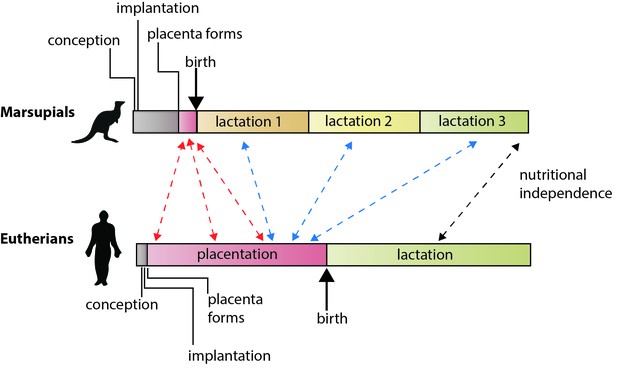
Az eutheri emlősök és az erszényes emlősök eltérő szaporodási stratégiái.
A eutheriákban az anya által a születés előtt (placentáció útján) és a születés után (laktáció útján) az utódok felnevelésébe fektetett energia nagyjából egyenlő mértékben. Az erszényesekben a vemhesség rövid, a placenta a vemhesség végén alakul ki, a laktáció pedig elhúzódik. Guernsey és munkatársai megmutatják, hogy az eutriákban a placentán keresztül történő fejlődést szabályozó genetikai jellemzők közösek a rövid életű erszényesek placentájával (piros nyilak). Azt is megmutatják, hogy az eutriákban a placenta funkcióit megalapozó gének némelyike kifejeződik a laktáció során az erszényesekben (kék nyilak), beleértve magának a laktációnak különböző konzervált összetevőit (fekete nyíl; Lefèvre et al., 2010). Megjegyzés: az időskálák nem abszolút értékek.
Az eLife-ban Julie Baker (Stanford University School of Medicine), Marilyn Renfree (University of Melbourne) és munkatársai – köztük Michael Guernsey (Stanford) első szerzőként, Edward Chuong (University of Utah) és Guillaume Cornelis (Stanford) – új részleteket közölnek az eutriák és erszényesek placentációjának és laktációjának hátterében álló molekuláris mechanizmusokról (Guernsey et al., 2017). Az eredményekhez az RNA-seq nevű technika módosított változatát használták annak mérésére, hogy a transzkriptom (a sejtben vagy sejtcsoportban lévő RNS-átiratok teljes készlete) hogyan változik a különböző sejttípusok között a fejlődés során (Rokas és Abbot, 2009).
Guernsey és munkatársai egy kis ausztrál erszényes, a tammar wallaby méhlepényének két sejttípusában a fejlődés során bekövetkező génexpressziós változásokat hasonlították össze. Azt találták, hogy a génexpresszió különbözött a két szövet között, ráadásul dinamikusan változott az idő múlásával, hasonlóan ahhoz, ami az eutriáknál történik. Ráadásul a transzkriptumok között sok olyat találtak, amelyeknek kritikus funkciójuk van az eutheri placentában, köztük az Ig7 jelátviteli útvonal tagjait és a GCM1 transzkripciós faktort, amely fontos a placenta kialakulásában és fejlődésében. És nem egyszerűen a gének konzerválódtak, a wallaby placentában a génexpressziós mintázatok hasonlítottak az egérplacentában a terhesség korai szakaszában megfigyeltekhez. Pontosan ezt várnánk, ha a méhlepény korai fejlődési funkciókat látna el a wallabyban, és a későbbi funkciókat a szülés után látná el. Ez arra utal, hogy az erszényesek és az eutriák közötti lényeges különbség nem a méhlepény korai funkcióiban van, hanem inkább abban, hogy a méhlepény funkciói hogyan tagolódtak az eutriák terhességének evolúciója során.
Végezetül Guernsey és munkatársai jellemezték a génexpressziós mintázatokat a tammar és több emlős emlőmirigyében. Mind az egér, mind a wallaby hasonló génexpressziós mintázatokat mutatott, ami aláhúzza a funkcionális kompartmentalizáció és konzerválás témáját mindkét csoportban. A legfigyelemreméltóbb azonban, hogy számos olyan gént azonosítottak, amelyek a tammar emlőmirigyeiben fejeződnek ki, és amelyekről ismert, hogy eutriákban a méhlepényben funkcionálisan fontosak (1. ábra). Ezek közé a gének közé tartoztak a tápanyagszállításban részt vevő gének és több olyan gén is, amelyekről ismert, hogy szükségesek az eutheri placentációhoz (köztük a GCM1). A génexpressziónak ez a konzerváltsága amellett érvel, hogy az erszényesekben a placenta a korai magzati fejlődést, a laktáció pedig a késői magzati fejlődést irányítja, ugyanazokat a géneket és molekuláris útvonalakat használva, mint az eutheri placenta.
Az erszényeseket tanulmányozók már régóta érvelnek amellett, hogy korrigálnunk kell tankönyveinket, hogy elismerjük az erszényes placentát és a sajátosan összetett laktációt (Renfree, 1983). Guernsey és munkatársai megerősítik ezt az érvelést annak bemutatásával, hogy mind az eutriák, mind az erszényesek konzervált gének eszköztárát fejezik ki, amelyek különböző szövetekhez és szervekhez lokalizálódhatnak, de közös célokat szolgálnak a magzati fejlődésben. Ez a meglepő konzerváltság aláhúzza az evolúció során bekövetkező funkcionális változások hátterében álló gének azonosításának fontosságát (Rausher és Delph, 2015).
A jövőre nézve érdemes megjegyezni, hogy az erszényesek óriási mértékben különböznek a reproduktív tulajdonságok tekintetében (Tyndale-Briscoe, 2005), és több faj jellemzése úgy, ahogyan azt Guernsey és munkatársai a tammar wallaby esetében megtették, gazdagabb képet fog nyújtani magának az erszényes vemhességnek az evolúciójáról és diverzitásáról. További munkára van azonban szükség a transzkriptom-profilok fajok közötti konzerváltságának számszerűsítésére alkalmas statisztikai módszerek kidolgozásához. És ha az emlősökön túlra tekintünk, a placentáció formái a gyíkoktól kezdve a csikóhalakon át a rovarokig mindenhol megtalálhatók, és az előzetes vizsgálatok azt jelzik, hogy az érintett gének vagy tulajdonságok közül sok közös (Ostrovsky et al., 2016; Whittington et al., 2015). Lenyűgöző lesz megtudni, hogy milyen mélyen tudjuk nyomon követni a terhességi eszköztár eredetét.