Introduction
メタン(CH4)は主要な温室効果ガスであります。 湿地、海洋、堆積物などの自然環境から排出されるメタンが、地球上の大気中のメタンの70%以上を占める(IPCC, 2007)。 公表されているデータを評価したところ、湿地、湖沼、河川、河口、海洋(順に補足図S1)のそれぞれで、メタン排出量が異なることがわかった。 自然の湿地だけで、生物起源CH4生成量の62%を占め(Kirschke et al. 一方、広大な面積を持つ海洋生態系は、CH4の自然発生源に対して約8%しか寄与していない(Nazaries et al.)
メタンの生物学的生成であるメタン生成は、メタン生成古細菌によって行われ、無酸素状態で主にH2/CO2、メチル基または酢酸からメタンを生成する(Thauer et al.、2008年)。 この反応はメチル-コエンザイムMレダクターゼ(MCR)によって触媒される。 この酵素のサブユニットをコードするmcrA遺伝子は、分子生物学的調査でよく用いられる遺伝子マーカーである(Conrad, 2007; Bridgham et al., 2013)。 mcrA遺伝子マーカーの利点は、メタン菌の系統的および機能的シグネチャーの両方を捕らえることであり、この特定の機能に対して高い配列決定深度を提供する(Lutonら、2002;Borrelら、2013;Yangら、2014)。 様々な自然環境から多数のmcrA配列が検索された。 公開されたmcrAデータセットにより、一般的な生態学的パターンを抽出し、地球規模や地域規模で形成される環境勾配を調査することが可能である。 さらに、152のメタン菌分離株の生理的特性をまとめたデータベースも公開されている1 (Jabłoński et al., 2015)。 最近、ゲノムビニングにより、Methanofastidiosaという新しいクラス(Nobu et al., 2016)やBathyarchaeota(Evans et al., 2015)、Verstraetearchaeota(Vanwonterghem et al., 2016)という新しい門に珍しいmcrA配列が発見された。 これらの新しい知見は,潜在的なメタン生成菌の多様性に関する我々の知識を拡大したが,大多数のメタン生成群集に対する分子マーカーとしてのmcrA遺伝子の適用性を不明瞭にするものではなかった。
現在までに,メタン生成群集は湿地,堆積物,永久凍土地帯,水田,消化器,地熱泉,熱水噴出孔で検出されている(Conrad,2007;Thauer他,2008;Wagner and Liebner,2009). メタン生成コミュニティ構造は,異なる空間・時間スケールで,環境pH,温度,塩分,地下水位,植生動態と関連していることが分かった(Megonigal et al., 2005; Milferstedt et al., 2010; Frank-Fahle et al., 2014; McCalley et al., 2014; Cui et al., 2015; Liebner et al., 2015)。 例えば、酢酸メタン生成は、一般的に低pHによって酢酸解離が低下するため妨げられる(Megonigalら, 2005; Kotsyurbenkoら, 2007)。 植物は、根からの滲出物やデトリタスの形で、メタン生成細菌の燃料となる可溶性の高品質有機炭素を供給できるため、主にフェンスでは、植物の滲出物が酢酸分解メタン生成細菌に有利となる(Bridgham et al.) 海水からの硫酸塩は潮間帯湿地でのメタン生成を抑制し、その結果、塩分濃度はメタン排出の一般的な予測因子として用いられてきた(Holm et al.、2016)。 チベットの湖沼堆積物に関する研究では、塩分濃度の上昇は水素栄養メタン生成菌を抑制するが、酢酸菌メタン生成は促進することが示された(Liu et al.、2016)。 これらの研究は、メタン生成コミュニティの環境ドライバーを示したが、単一の生息地または限られた空間スケールに焦点を当てている。
しかし、異なる環境変化に対するメタン生成細菌の適応を理解するには、微生物コミュニティ組成と環境条件との相関関係を体系的かつグローバルに調査する必要がある(Lozupone and Knight, 2007)。 現時点では、メタン生成生物群集の分散と生息域のフィルタリングについて取り上げた研究はわずかである(Auguet et al.2010; Barreto et al.2014)。 我々は、メタン生成生物群集は主に生息域のフィルタリングの影響を受け、それは地球環境の制御によって駆動されていると仮定する。 メタン排出速度が自然生態系間で大きく異なることを考慮すると、これらの生態系におけるメタン生成生物群集の組成、多様性および生物地理学を明示的に統合することは、現在および将来の気候変動に対するメタン生成の反応を決定するための基礎となる可能性がある。 本メタ研究は、メタン生成生物地理、多様性およびその環境制御に関するギャップを埋めるために、公開されているmcrA配列データおよび文献を用い、メタン生成分離株の生理学的データによって補完するために実施された。 各ヒットについて、原著論文を確認し、該当するmcrA配列をカスタムPerlスクリプトでパースした。 我々は自然環境に焦点を当てたので、メタン生成mcrA配列は自然生息地から取得し、土壌、湖沼、河口、海洋および熱水沈殿物、および泥火山に分類された。 クローンライブラリーの配列に加え、次世代シーケンス(NGS)から得られた5つのライブラリーを含む。 配列は、オリジナルデータセットにおける相対的な存在比を考慮せずにダウンロードした。 クローンライブラリーの配列は主に豊富な系統をカバーしているが、NGSはより深い多様性を捉えることができるため、シークエンス方法の解像度による誤差を考慮し、妥協してNGSのデータを使用することにした。 そこで、相対量が1%以上の豊富なOTUの代表的な配列のみを選択した。 さらに、塩基配列からタンパク質配列への翻訳チェックに失敗した配列や、低品質な配列(< 250 bps)は除外した。 最終的に、全世界に分布する94の部位から得られた4466のユニークなmcrA配列を含むデータセットを構築した(図1、補足表S1、S2)。 なお、ゲノム調査で必然的に検出される古細菌性メタン栄養細菌のmcrA配列は、データセットから差し引かなかった(Conrad, 2007)。 この部分は本研究の対象外である。
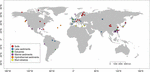
FIGURE 1. 本研究で使用したサイトの位置。 生息地によって6つのカテゴリーに分類し、異なる色でラベル付けを行った。 6つの生息地の概要は補足表S1、S2に示す。
本研究で検討した各調査地の地理座標とpH、塩分、標高、年平均気温(MAAT)、年平均降水量(MAP)などの環境設定は、データがある場合に対応する出版物から抽出した(補足表S1参照)。 複数の研究にまたがる環境パラメータの欠落を考慮し、関連文献のサイト記述に従っていくつかの環境変数を定性的に定義し、これらのカテゴリーデータを半計量的数値に変換した。例えば、海洋堆積物、熱水性堆積物、火山泥、ソーダ湖堆積物サンプルを「塩性」、土壌と淡水湖堆積物サンプルを「非塩性」、マングローブと河口のサンプルを「混合」サンプルとして定義している。
Raw Sequence Processing
シーケンス処理は、Mothurソフトウェアプラットフォームで実施した(Schloss et al, 2009). 異なるライブラリからの配列は、処理前にプールされた。 長さが350 bp未満の配列、または8塩基以上のあいまいな配列は破棄された。 その後、これらの配列を、FunGeneデータベースからhttp://fungene.cme.msu.edu/で検索したmcrA配列の事前整列サブセットに対して整列させた(Fish et al.、2013)。 キメラ配列は、データセット自体を参照として、Mothurソフトウェアでuchime法(Edgar et al., 2011)を用いて同定された。 次に、有効なmcrA遺伝子塩基配列を使用して、整列したDNA配列間の未補正ペアワイズ距離を計算し、さらに16S rRNA遺伝子の97%に相当する84%のカットオフで操作的分類単位(OTU)に割り当てた(Yang et al.、2014年)。 各mcrA OTUの存在量は、存在するかしないかのみで計上した。 ヌクレオチド配列とアミノ酸配列の両方を考慮することで、OTUの分類精度を向上させた。 DNAレベルでは、参照データベース(Yang et al.、2014)に従って、Mothurプラットフォームにより分類学的同一性を割り当てた。 タンパク質レベルでは、整列したタンパク質配列を用いてARBでツリーを構築し、対応するデータベースに基づいて分類学的な割り当てを行った。 OTUの割り当てが矛盾する場合は,NCBIで塩基配列とタンパク質配列の両方を手動でブラストし,クエリ・カバレッジ(>95%),同一性(>84%),E値(<1E-5)を考慮して,最終的に分類学上の同一性を決定した. タンパク質配列については、属レベルでのカットオフは83.5%の閾値を参照した(Hunger et al., 2011)。
Ecological and Statistical Analysis
統計解析には、各種Rパッケージを用いている。 主座標分析(PCoA)序列は、veganパッケージv2.2.0(Oksanen et al., 2015)を用いて構築したJaccard距離行列に基づいて生成された。 順列MANOVA(多変量分散分析)は、104の順列を持つビーガンのJaccard行列(McArdle and Anderson, 2001)の変動源を評価するために実施された。 Jaccard距離尺度は種の存否に基づいており、ほとんどの研究が代表的な配列のみを提供し、存在量に関する情報は欠落しているため、我々のデータセットにはより適している。 生息地ごとの分類学的出現頻度は、ggplot2パッケージ(v1.0.0)(Wickham, 2009)を用いたバブルプロットによって可視化された。 非塩水土壌および湖沼堆積物群集の階層的クラスタリング解析は,R関数「hclust」(R Core Team, 2014)により行った。 得られた群集クラスターは、元のサンプルのpHと温度レジームに従って記述した。これは、事前にPCoAを用いて、非塩水土壌と湖沼堆積物におけるメタン生成群集組成に影響を与えるパラメータとしてpHと温度の両方を同定したためである。 各メタン生成系統とこれらのクラスターとの関連性は、相関に基づく指標種分析(Dufrene and Legendre, 1997)を用いて決定された。 指標種は、特定の種類の生息地に豊富に存在し(特異性)、その種類の生息地で主に見られる(忠実性)種と定義される。 本研究では、非塩水土壌と湖沼堆積物について、指標種の概念と同様の指標分類群を、Rパッケージ labdsv (Roberts, 2016) による指標値(IndVal値)に従って、指定した反復にわたって観測値と同程度の指標値を得る確率が 0.05 未満であれば摘出することとした。 Chao2 指標は、vegan パッケージを使用して各サンプルについて計算した。 生息地間のChao2指標のWilcoxon順位和検定は、R関数「wilcox.test」(R Core Team, 2014)により実施した。 メタン生成古細菌分離株に対する生息地のフィルタリングの影響を集約するために、記載されたメタン生成培養物の生理学的および生化学的特性を、the3 ‘Methanogenic archaea database’ (Jabłoński et al., 2015) から取り出した。 その中で、最適なNaCl要求量の情報を持つ分離株をフィルタリングし、分離元に応じて分類・プロットした。
メタン菌の群集構造に対する分散制限の影響を調べるため、R関数「lm」(R Core Team, 2014)により地理的距離行列と群集Jaccard距離行列に基づいて線形回帰分析が行われた。 Rのveganパッケージ(Oksanen et al., 2015)を用いて、再度2つの行列に応じた分散制限の効果を評価するために、Mantel検定および部分Mantel検定を実施した。 さらに、R ade4パッケージの関数「multispati」(Dray and Dufour, 2007)により、メタン菌の空間構造を探るために、モランのI指数に基づく多変量空間分析(空間PCA)を16のヨーロッパの土壌および湖底堆積物サンプルに適用した。 さらに、これらの16サンプルに対して、R関数「hclust」(R Core Team, 2014)を用いて、Jaccard距離行列に基づくWardの最小分散クラスタリングを実施し、さらにクラスタリングの結果を地理地図上に投影した。 州レベルでマッピングするための欧州のシェープファイルは、GSHHG Database (v2.3.6)4 で入手可能である。 地図はQGIS v2.18.25で作成した。
結果
自然環境におけるメタン生成アーキアの生物地理
世界に分布する94の自然環境からmcrA遺伝子配列を検索した。 これら94地点のそれぞれの位置と生態系の種類を図1に示す。 メタン生成系統の発生(有無)頻度を生態系の種類に応じてマージし、図2に示した。 土壌ではMethanoregulaが最も多く、Methanobacterium、Methanosaeta、Methanocella、Methanomassiliicoccus、Methanosarcinaと一緒に観察されることが多い。 河口堆積物では、Methanosaeta、Methanobacterium、Methanoregula、Methanoculleusの塩基配列がよく検出された。 また、湖沼堆積物では、MethanoregulaとMethanosaetaが主に検出された。 海底堆積物では,MethanoculleusとMethanosaetaが最も多く,次いでMethanolineaであった。 異なる自然環境におけるメタン生成菌系統の発生頻度を示すバブルプロット。 縦軸に沿った順位は、系統の総発生頻度の減少に対応している。 分類は属レベルで示した。 属レベルでの分類が不可能な場合は、次に高い分類レベルが使用された。 5719>
多くの分類群が異なる環境で検出されたにもかかわらず、環境選好性を示すものがある。 Methanoregulaは非海洋および過渡的環境(土壌、湖沼堆積物、河口)で頻繁に出現する分類群であり、海洋生息地(海洋堆積物、熱水堆積物、泥火山)には存在しない。 メタノバクテリウムとメタノセラは、非海洋および過渡的な環境で優勢であるが、海洋の生息地ではほとんど見いだせない。 一方、Methanococcoidesは、海洋堆積物、熱水堆積物、泥火山で優勢な系統であるが、土壌や湖沼堆積物ではほとんど観察されない。 さらに、MethanogeniumとMethanolaciniaは河口や海洋堆積物にのみ存在し、MethanospirillumとMethanosphaerulaは陸上環境のみに存在する。 また、Methanocaldococcus、Methanothermococcus、Methanopyrus、Methanotorris、Methanococcusなど、熱水系堆積物にのみ存在する分類群もある。 メタノサエタなど一部の系統はほとんどの環境に存在するが,遍在する系統はない。
系統が最も豊富なのは河口堆積物で,様々な系統の出現頻度もより均等であった。 一方、泥火山や熱水生態系では、メタン生成菌の多様性は相対的に低いことが示された。
Alpha-Diversity of Methanogenic Communities in Natural Environments
Chao2 indexによるメタン生成アーキアの豊かさは、生態系のタイプによって大きく異なる(図3)。 アルファ多様性を直接比較し、試料間の合理的なトレードオフを得る目的で、各サイトについて15配列へのサブサンプリングを行った。 Chao2指標によると、河口堆積物は6つの生態系タイプの中で最もメタン生成菌の種が豊富であり(補足表S3)、これはバブルプロットの結果(図2)を裏付けている。 土壌と湖沼堆積物は、河口域のサンプルよりも低い豊かさを示したが、海洋堆積物や熱水性堆積物よりも有意に高いChao2指数を示し、海洋堆積物と熱水性堆積物の間には有意差はなかった(補足表S3)。 このプロットは、アルファ多様性指標での比較をより強固にするために、各サイトの15配列を含むサブサンプルデータセットに基づいている。 各生息地のサンプル数は、生息地のラベルの下に’n’として示されている。 ns’はWilcoxon検定で統計的に有意でないことを示す。 OTUレベルのα多様性の統計結果はSupplement Table S3に示した。
自然環境におけるメタン生成生物群集のグローバルコントロール
世界的に分布する94のメタン生成生物群集をJaccard距離行列に基づくPCoAを用いて序列プロットにクラスタリングした。 PCoA解析の結果、第1軸と第2軸を合わせて全分散の16.3%を説明することができた。 このことから、試料間のばらつきは、塩分によってほぼ説明できることがわかった(図4)。 塩分濃度の初期データが得られない場合もあったため、これらの試料を上記のように塩分、混合(中間)、非塩分の各試料に定性的に分類した。 塩水性試料と非塩水性試料は、第一軸に沿って効果的に分離している。 混合試料は、全体として生理食塩水と非生理食塩水の中間的なグループになる。 ジャカード距離行列に基づく並べ替えMANOVAは、塩分が世界のメタン生成コミュニティの分布を制御する主要な生物学的因子であることも示唆している(R2 = 0.099, P < 0.001)(Table 1)。 94サンプルを比較したメタン菌mcrA遺伝子配列ライブラリのJaccard距離行列に基づく主座標分析(PCoA)序列化。 PCoAは塩分濃度によって色分けされている。 赤色は非塩分環境、青色は塩分環境、緑色は中間的な環境を示す。 異なるシンボルは異なる環境を表す。 表1

TABLE 1.主座標で説明される変動の割合を軸に示す。 コミュニティ分散の異なる環境変数との関連性を検証するために、6つの生息地の全サンプルのJaccard距離行列に対するパーミュテーションMANOVA分析。
さらに、メタン生成純粋培養の分離源と増殖に最適なNaClの濃度との間の関係の可能性について確認した。 メタン生成純培養物の最適なNaCl濃度は、海洋から河口、湖沼堆積物、土壌生態系へと減少することが示された(補足図S2)。
Environmental Controls and Methanogenic Indicator Taxa in Non-saline Soils and Lake Sediments
地球規模で見ると、非塩性土壌と湖底堆積物のメタン生成コミュニティは密接にクラスター化している(図4)ため、合計33の調査サイトを占めるこれら2つの生息地のメタン生成コミュニティの環境制御についてさらに分析を行った。 この2種類の生息環境について、群集ベースのクラスター分析を行った結果、Jaccard距離行列に基づく4つのクラスターが存在した(補足図S3)。 並列MANOVAは、pH (R2 = 0.099, P < 0.001) と温度 (R2 = 0.069, P < 0.001) が非塩性土壌と湖底堆積物のメタン生成β多様性に影響を与えることを示唆している(表2)。 そこで、4つのクラスターを最初のサンプリング地点のpHとMAATに割り当てたところ、群集のクラスタリングにほぼ一致するサブグループが得られた(図5C,D)。 環境特性とこれら4つの群集クラスターの組み合わせにより、これら4つのサブグループを中性・低温グループ1、酸性・低温グループ2、酸性・中程度グループ3、中性・温暖グループ4と定義することができる(図5)。 さらに、Jaccard非類似度行列に基づくPCoA順序付けにより、PC1軸に沿って、グループ1とグループ4が主に中性環境であるのに対し、グループ2とグループ3の試料の多くは酸性土壌と湖底堆積物から得られることが示唆された(図5A)。 さらに、PC2軸に沿って、中程度の場所(グループ3)のサンプルは温暖な場所や寒冷な場所のサンプルと分離し、一方、PC3軸に沿って、温暖な環境(グループ4)のサンプルは他のサンプルと分離している(図5B)。 このように、PCoA順序付けの最初の3軸は、全変動の38.8%を説明する。

TABLE 2. また、非塩水土壌と湖底堆積物サンプルのJaccard距離行列に基づく並べ替えMANOVAにより、異なる環境変数と群集分散の関連性を検証した。 33種類の非塩水性土壌と湖沼堆積物に対するジャカード距離行列に基づく主座標分析プロット。 サブプロット(A)はPC1とPC2を示し、記号はpHで着色、サブプロット(B)はPC2とPC3を示し、記号は温度で着色している。 最初の3成分は分散の17.1、11.7、10%を説明する。 箱ひげ図は、4つのサブグループのpH (C) とMAAT (年平均気温) (D)を示したものである。 図Cの箱の色は(A)のpHの区分に対応している。 箱ひげ図の色は、ペアワイズ・ウィルコクソン検定 (P < 0.05) に基づく統計的有意性を示しており、同じ色のサンプルは互いに有意な差はないことを示している。 同様に、(D)の色は(B)の温度グループ分けに準じている。 サブグループは群集の類似性の階層的クラスター分析を参照する。
発生頻度に基づいて、各サブグループにおけるメタン生成系統の発生を調べた(図6)。 Methanoregulaはすべてのタイプの非塩水環境において優勢であった。 また,中性・低温サブグループ(グループ1)では,Methanoregulaに加えて,Methanosaeta,Methanobacterium,Methanosarcinaが多く出現した. 酸性・寒冷域のグループ2には、Methanobacterium、Methanocella、Methanosarcinaが含まれ、Methanosaetaは含まれない。 酸性・中温性グループ3ではMethanocellaとMethanosaetaが多く、中性・温熱性グループ4ではMethanolineaとMethanosaetaが重要なメンバーである。 このグループはMethanoculleusが確認された唯一のグループである。

FIGURE 6. 土壌と湖底堆積物のメタン生成生物群集の4定義グループ内のメタン生成系統の発生頻度. 縦軸はアルファベット順に配列されている。 棒の長さは、対応するグループ内の各系統の平均発生頻度に相当する。 エラーバーは、そのグループ内の異なるサンプルに対するある分類群の標準偏差を表す。 アスタリスクはP-value < 0.05の専門家分類を示す。 分類学は属レベルのものを示す。 属レベルの分類が不可能な場合は、次に高いレベルの分類を使用した。 図5に4グループの説明を示す。
4グループとも、発生率の高い分類群は、Methanoregula、Methanobacterium、Methanosarcina、Methanosaeta、Methanomassiliicoccus、Methanocellaであった。 特定のグループ内のほとんどのサイトに有意に多く存在する専門分類群は、前述したように指標種分析に従って検出された。 その結果、31分類群のうち6分類群が有意な指標値(P < 0.05)を示した(図6にアスタリスクのラベルを付した)。 グループ1(中性・寒冷)では、Methanosaeta、Methanolobus、Methanomethylovoransの系統が最も多く、専門家が多く存在した。 Methanobacteriumは寒冷・酸性域の第2グループで、Methanolineaは第4グループ(中性・温暖)でスペシャリストとして確認されたが、他のグループにはほとんど観察されなかった。 また、Methanoregulaは酸性・中性のグループ3に多く存在した。
分散制限
線形回帰分析(R2= 0.05, P < 0.001)は、グローバルデータセットにおいて地理距離とメタン生成コミュニティ構造の間に弱い相関があることを示唆した。 同時に、マンテル検定により、環境変数が地理的距離よりも群集構造に対して高い相関を持つことが示された(表3参照)。 この傾向は、自己相関の影響を制御した偏マンテル検定でも確認された。 地理的距離とJaccard群集類似度をプロットすると、明確な線形傾向は見られないが、主にサンプリング地点の世界的な分布に起因するパターンが見られた(補足図S4参照)。 Mantelおよび部分Mantel検定分析による、グローバルデータセットおよびヨーロッパ16サンプルのサブサンプルの微生物分布に対する環境変数および地理的距離の影響の決定。
分散性の影響をさらに分析するために、最も密で均一にサンプリングされたヨーロッパに分析を限定しました。 この部分集合に対するMantel検定および部分Mantel検定では,群集データは地理的距離よりも環境変数に高い相関があるという傾向が再現された(表3参照)。 環境変数を制御した部分マンテル検定では、微生物群集と地理的距離の間に統計的に有意な相関は検出されなかった。 これら 16 のヨーロッパのサイトに関する空間 PCA 分析は、メタン生成コミュニティの空間構造(全分散の 23.7% がこの構造で説明される)を示唆し、これはモランの I 指数(モランの I = 0.4018) によって示されるサイトの正の空間自己相関に相当する。 第一固有値のみが安定で、中央ヨーロッパとバルト三国の間のサンプルの分離に対応した(補足図S5)。 しかし、データセットが小さいため、この観察された空間構造を地理的変数、環境変数、またはその両方にしっかりと割り当てることは困難である。 そこで、上記のようにメタン生成生物群集のクラスター分析を行い、3つのグループ(図7A)を発見し、地理的な地図に投影した(図7B)。 このクラスタリングは、バルト海に沿った空間PCAの分離を再現していなかった。 したがって、地理的に非常に近いサイトでも、異なるメタン生成生物群集構造を示し、異なるクラスタに組み立てられていることがわかった。 一方、地理的に非常に離れた地点では、非常に類似した群集組成を示し、クラスターが形成されている(図7B)。 ヨーロッパにおけるグループの地域的な分散は、地理的な構造に対応するものでも、それによって制限されるものでもないようである
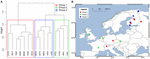
FIGURE 7. ヨーロッパの土壌と湖底堆積物16試料間のJaccard距離行列の階層的クラスター分析。 解析にはウォードクラスタリング法を用いた。 クラスタリングデンドログラム(A)では異なる長方形で色分けされた3つのグループに分けられ、ヨーロッパ地図(B)に投影された。
議論
微生物群に対する生物地理学の概念の特定と適用は、微生物生態学の大きな関心事である。 微生物の生物地理学は、生息地のフィルタリング、分散、ドリフト、突然変異という4つの主要なプロセスの進化的・生態的相互作用によって支配されていると考えられている(Hanson et al.、2012)。 ドリフトと突然変異の影響は本研究の解釈力を超えているが、我々は自然環境におけるメタン生成生物群集のグローバルなパターンが存在することを示す。 本研究では、メタン生成生物群集のグローバルな生物地理学的パターンが、地理的分散よりも生息地のフィルタリングと関連していることを実証した。 例えば、ソーダ湖堆積物のメタン生成生物群集は、地理的に離れた海洋試料と密接にクラスター形成しており(図4)、ヨーロッパの土壌と湖堆積物には、互いに大きく離れた場所にあるにもかかわらず、非常に類似したメタン生成生物群集が存在している(図7)。 全体として、分散制限の可能性を明らかにするための我々の試みは、メタン生成生物群集構造に対する地理的位置の影響は弱く、環境条件の明確な影響とは対照的であった。 微生物群集に対する空間的な影響を決定的に排除することは、利用可能なデータポイントでは不可能である。 サンプリング地点は特定の生息地や地域に集中しており、全体のサンプル数は少ない。 もし影響があるとすれば、空間的な影響は地域的または局所的なものだけであると考えられる。 超好熱性古細菌では地理的障壁による局所的な分散制限が報告されており(Whitaker et al., 2003)、アンモニア酸化細菌では地域・大陸スケールではなく局所的な塩湿地での分散制限が報告されている(Martiny et al.2009)、そして、ほとんどがアロキソン性炭素の利用可能性によって引き起こされる大規模な不均質な雪環境の細菌に対して(Lutz et al.) 自然環境におけるメタン菌に対する分散制限の影響が小さいということは、メタン菌が空間上にランダムに分布し、その生理的要件が満たされれば地域環境によってうまく選択され、安定した群集を確立できることを意味する(Martiny et al.2006; von Mering et al.2007)
生息地フィルタリングという概念は、同様の生態的要件を備えた種が偶然に予想されるよりも多く共起するはずだということを意味している(Weiher and Keddy, 2001; Cornwell et al.2006; Ulrich et al.2010). その結果、海洋生態系と湿地・湖沼生態系の間でメタン生成生物群集組成に大きな差が生じ、河口はその中間に位置することが明らかになった。 環境サンプルの一連の遺伝子調査に基づく生物地理学的パターンは、一般細菌(Nemergut et al., 2011)、メタン湧出微生物群(Ruff et al., 2015)、アンモニア酸化古細菌(Cao et al., 2013)、海洋外洋・底生細菌(Zinger et al., 2011)、窒素循環微生物(Church et al., 2008)でも報告されている。 生息域のフィルタリングは、未培養古細菌(Auguet et al., 2010)、多様な環境における細菌群集全体(von Mering et al., 2007; Chaffron et al., 2010)または南米の泥炭地における地域スケールで具体的に報告されている(Oloo et al, 2016)、メタン酸化細菌(Knief, 2015)や窒素固定細菌(Nelson et al., 2016)といった特定の細菌群も含まれている。
我々の結果は、地球規模では塩分がメタン生成群集の構成を大幅に制御し、海洋と陸上のメタン生成群集の大きな違いを決定していることを示した。 また,ソーダ湖堆積物のメタン生成菌は海洋堆積物のメタン生成菌とクラスター化し(図4),塩分濃度のグローバルな影響が強調された。 この結果は、塩分濃度が細菌群集および古細菌群集のグローバルなパターンを形成する主要な要因であるとする16S rRNA遺伝子に基づく他の研究(Lozupone and Knight, 2007; Auguet et al, 2010; Caporaso et al, 2011; Cao et al, 2013)と同様である。 地理的分離の影響は低いが、塩分の影響が一般的な微生物群集に強いことも、先行研究で観察された(Logares et al., 2013; Yang et al., 2016)。 したがって、塩分濃度は、どの系統が生き残れるかを大きく左右する。 様々な生息地でメタン生成活性は塩分と負の相関があった(Bartlett et al., 1987; Potter et al., 2009; Poffenbarger et al., 2011)。 そのため、塩分によるメタン生成の阻害は、メタン生成菌の個体数の減少と一致することが示唆されている(Pattnaik et al.) 水素栄養、酢酸栄養、メチル栄養のメタン生成に対する塩分の効果は、塩分のレベルに依存し、メタン生成の異なる経路で異なる(Liu et al.) 現在のところ、塩分が群集構造に与える影響を説明する明確なメカニズムはないが、いくつかの仮説が説明の可能性として考えられる。 生理的には、塩分濃度は細胞の外部および内部の浸透圧に影響を与える。 非塩性メタン生成細胞は、内部浸透圧に対抗するための生理的適応を発達させているが、塩適応細胞はそのような機能を失っている (Zinder, 1993)。 さらに、塩分濃度が上昇すると、メタン生成細胞は、適合溶質の合成や取り込みの比率を高め、エネルギー的、代謝的に大きな犠牲を強いられることになる(McGenity, 2010)。 海底堆積物や熱水堆積物から分離されたメタン生成菌は、土壌から分離された菌よりも最適なNaCl濃度が有意に高いことがわかったため、耐塩性の特性はメタン生成菌の純粋培養の最適濃度にも現れている(補足図S2)。 したがって、これらの環境のメタン生成細菌は4つのグループに分類される(図6)。 海洋生態系とは異なり、非塩分の陸上生態系は、pHと温度の両方に大きな自然変動がある。 温度は、メタン生成経路だけでなく、メタン生成菌の集団自体にも影響を与える(Conrad、2007;Rooney-Varga et al.、2007)。 発酵と酢酸発酵の過程で温度に敏感な段階があるため、温度が上昇するとメタン生成量が大幅に増加する可能性がある(Megonigal ら, 2005; Kotsyurbenko ら, 2007)。 さらに、低pHは、酢酸の解離を妨げることによって酢酸の利用可能性を大幅に制限し、酢酸メタン生成に負の影響を与えることがある(Fukuzaki et al.) このことは、Methanosaetaがgroup2には存在せず、Methanosarcinaは異なる供給源間で切り替えが可能で、大きな影響を受けなかった理由である可能性がある。 さらに、pHは、メタン生成経路の両方を直接的に阻害し、発酵への影響を通じて間接的に、オンブロトロフィックからミネラトロフィックな泥炭地のメタン生成効率とメタン生成経路を調節できる(Ye et al.、2012)。 したがって、温度とpHの両方が、メタン生成に関連する代謝ステップと、メタン生成菌に基質を提供する上流の発酵を直接的または間接的に調節する可能性がある。
メタノレギュラは4群の陸上生息地のすべてで遍在し非常に豊富だが(図6)、海洋系ではほとんど見られず、海洋領域の淡水の影響に関する代理人と見なされる可能性がある。 その世界的な関連性は、最近別のところで報告された(Yang et al.、2017)。 土壌や湖沼堆積物に遍在しているにもかかわらず、Methanoregulaは中程度の温度を持つ酸性の生息地で指標系統として発生する。 さらに、(1)Methanolineaは温暖で中性の陸上環境、(2)Methanobacteriumは低温で酸性の環境、(3)MethanosaetaはpH中性環境に特に適応しているようで、これは他の研究(Rosenberg et al,2014)と一致し、我々のアプローチの堅牢性を裏付けている。 一般に,メタン生成生物群集を取り巻く地球化学的条件は,ニッチ分化をもたらす. ニッチの選別は適応的な専門家を残す傾向があるため(Langenheder and Székely, 2011),長期的な環境選択の進行により,他の異なる環境条件では代表性が低いか存在しない可能性のある固有の生息地専門家の配列によって満たされた多様なニッチが生成された. また、生物群集は、生態学的相互作用、ダイナミクス、競争、共生などの生物学的要因によって形成される。 それらの生物学的要因にもかかわらず、von Meringら(2007)は、生息地の嗜好性はしばしば経時的に著しく安定しており、ひいては環境コミュニティの特徴的な分類群構成は、その生態と機能の重要な指標となり得ることを見出した。
メタン生成アーキアの生息地の嗜好性と一致して、近縁のメタン生成株はしばしば同等の環境から分離されていると思われた。 例えば,Methanoregulaceaeは自然環境ではかなり多様であるようで(Yang et al., 2017),中立的な株はこれまで培養に抵抗することがあった。 現在記載されているMethanoregulaの株はいずれもやや好酸性の環境から得られており、Methanolineaの2つの代表はそれぞれ消化器汚泥や水田土壌などの比較的暖かい生息環境から得られている(Rosenberg et al.、2014年)。 Methanobacteriumは様々な環境から分離されたが、本属の既存分離株の約半数は7よりやや低いpH至適を示す。このことは、環境配列に基づいて同定された指標系統が、既存のメタン生成培養菌の生理や供給源の分化を反映している可能性を示唆している。 例えば、生息地の一般的な特性である塩分濃度は、生物を強い環境選択に徐々にさらし、周囲の塩分濃度に最適な新しい種の集合をフィルターすることができる(Logares et al, 2013)。 ここでは、自然環境のメタン生成アーキアは、河口堆積物において最も多様であることを示す。 河口域は海洋生態系と陸上生態系の移行帯である。 このため、種の豊富さに寄与しうる2つの主要なプロセスが存在する。 第一に、海と陸の微生物が河口で混ざり合い、最終的に全体的に高い多様性を包含する(McLusky and Elliott, 2004)。 例えば、河口域のバクテリア、古細菌、真菌、さらにはユニークな機能を果たす特定のバクテリアの高い多様性が観察されている(Cunliffe et al.、2008;Mosier and Francis、2008;Crump et al.、2012)。 また、河口域の生物が餌とする陸域や潮流の入力により、栄養レベルが高いという側面もある(McLusky and Elliott, 2004; Statham, 2012)。 このような背景から、河口環境はメタン菌の一般的な新規性を回復させる上で重要である。 これまで、種の多様性が生態系プロセスに及ぼす影響については、相当な研究努力が払われてきた。 生物多様性と生態系機能の関連性はまだ議論中であり、微生物群集についてはまだ解明されていない(Loreau et al.、2001;Tilman et al.、2014)。 土壌や湖沼堆積物はメタンの主要供給源であり、またメタン生成の多様性が高い生息地でもあるが、我々は種の豊富さはメタン生成能や生態系メタン排出の適切な代理ではなく、むしろ環境の不均質性と歴史を反映していると思われることを提唱している。 種の豊富さによって環境をランク付けすることは、土壌や湖から最も多く、河口からは比較的少ないメタン排出率の可能性を必ずしも意味しない(図3および補足図S1)。 文献や公開データベースでメタン菌に関する配列データが増えているにもかかわらず、提供される関連する生体変数は一貫性がなく、まばらであることが多い。 環境変数に関する一貫した情報の量が限られているため、多変量統計解析の適用に制約がある。 本研究における生物学的要因は、群集の分散のごく一部しか説明できないことを考えると、他の説明変数が欠けていることが示唆される。 特に重要なのは、酢酸、水素、メチルアミンなどのメタン生成基質の濃度と利用可能性であろう。 しかし、生息地はそれぞれ異なる歴史を持ち、局所的にしか調査できないため、メタン生成生物群集の構造を完全に説明するには、生物学的パラメータだけでは十分でない可能性がある。 また、ロシアやカナダの亜寒帯や北極の広大な地域は、利用可能なmcrAデータの中で、十分に表現されていない。 また、ロシアやカナダの亜寒帯・北極圏の広大な地域では、mcrA遺伝子のデータが不足している。 XWとSYはデータの収集と解析を行った。 統計解析はXW、SY、FHが行った。 XW、MW、SYは系統樹の補正を行った。 XW、SY、FH、MW、DW、SLは結果を解釈し、論文を執筆した。
利益相反声明
著者らは、本研究が利益相反の可能性があると解釈される商業的または金銭的関係がない状態で行われたことを宣言する。
謝辞
中国奨学会によるXWへの財政支援(XWへの助成番号201408620031)については、感謝する意を表した。 本研究は、Helmholtz Gemeinschaft(HGF)からSLのHelmholtz Young Investigators Group(VH-NG-919)に資金援助を受けた。
補足資料
この論文の補足資料は、オンラインで見ることができます。 https://www.frontiersin.org/article/10.3389/fmicb.2017.01339/full#supplementary-material
脚注
- ^ http://metanogen.biotech.uni.wroc.pl/
- ^ http://www.ncbi.nlm.nih.gov/
- ^ http://metanogen.biotech.uni.wroc.pl/
- ^ https://www.ngdc.noaa.gov/mgg/shorelines/gshhs.html
- ^ http://qgis.osgeo.org
Auguet, J. C….., Barberan, A., and Casamayor, E. O. (2010). 未培養古細菌のグローバルな生態パターン. ISME J. 4, 182-190. doi: 10.1038/ismej.2009.109
PubMed Abstract |Ref Full Text | Google Scholar
Bange, H. W., Bartell, U. H., Rapsomanikis, S., and Andreae, M. O. (1994). バルト海と北海におけるメタンと,メタンの海洋排出の再評価. Glob. Biogeochem. Cycles 8, 465-480. doi: 10.1029/94GB02181
CrossRef Full Text | Google Scholar
Barreto, D. P., Conrad, R., Klose, M., Claus, P., and Enrich-Prast, A. (2014). 熱帯の湖沼堆積物におけるバクテリア、アーキア、メタン生成アーキアの距離減衰と分類群面積の関係。 PLoS ONE 9:e110128. doi: 10.1371/journal.pone.0110128
PubMed Abstract | CrossRef Full Text | Google Scholar
Bartlett, K. B., Bartlett, D. S., Harriss, R. C., and Sebacher, D. I. (1987). 塩性湿地の塩分濃度勾配に沿ったメタン排出量. doi: 10.1007/BF02187365
CrossRef Full Text | Google Scholar
Borrel, G., O’Toole, P. W., Harris, H. M. B., Peyret, P., Brugere, J. F., and Gribaldo, S. (2013). メチロトロフィックメタン生成菌の7番目の順番を支持する系統樹データ、およびメタン生成の進化に関する洞察を提供する。 Genome Biol. Evol. 5, 1769-1780. doi: 10.1093/Gbe/Evt128
PubMed Abstract | CrossRef Full Text | Google Scholar
Bousquet, P., Ciais, P., Miller, J., Dlugokencky, E., Hauglustaine, D., Prigent, C.、他 (2006).P. (2006). 大気中のメタン変動に対する人為的および自然起源の寄与。 Nature 443, 439-443. doi: 10.1038/nature05132
PubMed Abstract | CrossRef Full Text | Google Scholar
Bridgham, S. D., Cadillo-Kuiloz, H., Keller, J. K., and Zhuang, Q. (2013). 湿地からのメタン排出:局所から地球規模までの生物地球化学的、微生物的、およびモデリング的視点。 Glob. Chang. 19, 1325-1346. doi: 10.1111/gcb.12131
PubMed Abstract | CrossRef Full Text | Google Scholar
Cao, H., Auguet, J.-C., and Gu, J.-D. (2013). アンモニア酸化性古細菌のグローバルな生態パターン。 PLoS ONE 8:e52853. doi: 10.1371/journal.pone.0052853
PubMed Abstract | CrossRef Full Text | Google Scholar
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Lozupone, C. A., Turnbaugh, P. J.、他 (2011)(Camera et al. サンプルあたり数百万配列の深さでの16S rRNA多様性のグローバルパターン。 Proc. Natl.Acad. Sci. U.S.A. 108, 4516-4522. doi: 10.1073/pnas.1000080107
PubMed Abstract | CrossRef Full Text | Google Scholar
Chaffron, S., Rehrauer, H. , Pernthaler, J. , and von Mering, C. (2010). 環境と全ゲノム配列データからの共存する微生物のグローバルネットワーク。 Genome Res. 20, 947-959. doi: 10.1101/gr.104521.109
PubMed Abstract | CrossRef Full Text | Google Scholar
Church, M.J., Bjorkman, K.M., Karl, D.M., Saito, M.A., and Zehr, J.P. (2008). 太平洋における窒素固定菌の地域分布. Limnol. Oceanogr. 53, 63-77. doi: 10.4319/lo.2008.53.1.0063
CrossRef Full Text | Google Scholar
Conrad, R. (2007). 「メタン生成菌とメタン栄養菌の微生物生態”, Advances in Agronomy, ed. L. S. Donald (Cambridge, MA: Academic Press), 1-63.
Google Scholar
Cornwell, W.K., Schwilk, D.W., and Ackerly, D.D. (2006). 棲息地フィルタリングのための特性ベースのテスト:凸包体積。 Ecology 87, 1465-1471. doi: 10.1890/0012-9658(2006)872.0.CO;2
PubMed Abstract | CrossRef Full Text | Google Scholar
Crump, B. C., Amaral-Zettler, L. A., and Kling, G. W. (2012). 北極圏の淡水における微生物の多様性は、土壌からの微生物の接種によって構造化される。 ISME J. 6, 1629-1639. doi: 10.1038/ismej.2012.9
PubMed Abstract | CrossRef Full Text | Google Scholar
Cui,M., Ma,A., Qi,H., Zhuang,X., Zhuang,G., and Zhao,G. (2015). 気温の上昇は,メタン生成生物群集組成を変化させることなく,チベット高原のゾイゲ湿地からのメタン排出を加速させる。 Sci. Rep. 5:11616. doi: 10.1038/srep11616
PubMed Abstract | CrossRef Full Text | Google Scholar
Cunliffe, M., Schafer, H., Harrison, E., Cleave, S., Upstill-Goddard, R., and Murrell, J. C. (2008). 河口域の表面マイクロ層に生息する細菌および古細菌群集の系統・機能遺伝子解析 ISME J. 2, 776-789. doi: 10.1038/ismej.2008.28
PubMed Abstract | CrossRef Full Text | Google Scholar
Dray, S., and Dufour, A. B. (2007). ade4 パッケージ:生態学者のための二元性ダイアグラムの実装。 J. Stat. Softw. 22, 1-20. doi: 10.18637/jss.v022.i04
CrossRef Full Text | Google Scholar
Dufrene, M., and Legendre, P. (1997). 種集合と指標種:柔軟な非対称アプローチの必要性. Ecol. Monogr. 67, 345-366. doi: 10.2307/2963459
CrossRef Full Text | Google Scholar
Edgar, R. C., Haas, B. J., Clemente, J. C. , Quince, C., and Knight, R. (2011). UCHIMEはキメラ検出の感度と速度を向上させた。 Bioinformatics 27, 2194-2200. doi: 10.1093/bioinformatics/btr381
PubMed Abstract | CrossRef Full Text | Google Scholar
Evans, P.N., Parks, D.H., Chadwick, G. L., Robbins, S.J., Orphan, V. J., Golding, S. D., et al(2015).キメラ検出の感度と速度を向上させた。 ゲノム中心メタゲノミクスによって明らかになった古細菌門Bathyarchaeotaのメタン代謝。 Science 350, 434-438. doi: 10.1126/science.aac7745
PubMed Abstract | CrossRef Full Text | Google Scholar
Fish, J. A., Chai, B., Wang, Q., Sun, Y., Brown, C.T., Tiedje, J. M.、他 (2013). FunGene: 機能的遺伝子パイプラインとリポジトリ. フロント. Microbiol. 4:291. doi: 10.3389/fmicb.2013.00291
PubMed Abstract | CrossRef Full Text | Google Scholar
Frank-Fahle, B. A., Yergeau, E., Greer, C. W., Lantuit, H., and Wagner, D. (2014). カナダ北西部北極圏の永久凍土の影響を受けた土壌における微生物機能ポテンシャルと群集組成. PLoS ONE 9:e84761. doi: 10.1371/journal.pone.0084761.g001
PubMed Abstract | CrossRef Full Text | Google Scholar
福崎慎太郎・西尾伸夫・長井慎太郎 (1990). 酢酸のメタン発酵のカイネティクス. Appl. Environ. Microbiol. 56, 3158-3163.
Google Scholar
Hanson, C. A., Fuhrman, J. A., Horner-Devine, M. C., and Martiny, J. B. (2012). 生物地理学的パターンを超えて:微生物景観を形成するプロセス。 Nat. Rev. Microbiol. 10, 497-506. doi: 10.1038/Nrmicro2795
PubMed Abstract | CrossRef Full Text | Google Scholar
Holm, G. O., Perez, B. C., McWhorter, D. E., K. W., Johnson, D. J., Raynie, R. C., et al(2016).をご参照ください。 ミシシッピ川デルタの潮間帯の淡水・汽水湿地からの生態系レベルのメタンフラックス:沿岸湿地の炭素プロジェクトに対する示唆。 Wetlands 36, 401-413. doi: 10.1007/s13157-016-0746-7
CrossRef Full Text | Google Scholar
Hunger, S., Schmidt, O. , Hilgarth, M. , Horn, M. A. , Kolb, S. , Conrad, R., and al. (2011). 無酸素状態のメタン排出フェン土壌におけるギ酸および二酸化炭素資化性原核生物の競合。 Appl. Environ. Microbiol. 77, 3773-3785. doi: 10.1128/AEM.00282-11
PubMed Abstract | CrossRef Full Text | Google Scholar
IPCC (2007). “Climate Change 2007: the Physical Science Basis,” in Proceedings of the Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, eds D. Qin, S. Solomon, M. Manning, Z. Chen, M. Marquis, K. B. Averyt, et al. (Cambridge: Cambridge University Press).
Google Scholar
Jabłoński, S…, Rodowicz, P., and Łukaszewicz, M. (2015). 生理・生化学的特徴を含むメタン生成古細菌データベース。 Int. J. Syst. Evol. Microbiol. 65, 1360-1368. doi: 10.1099/ijs.0.000065
PubMed Abstract | CrossRef Full Text | Google Scholar
Kirschke, S., Bousquet, P., Ciais, P., Saunois, M., Canadell, J.G., Dlugokencky, E. J.、他 (2013).メタン菌の生理・生化学的特徴を含むメタン菌データベース。 30年にわたる世界のメタン発生源と吸収源。 Nat. Geosci. 6, 813-823. doi: 10.1038/Ngeo1955
CrossRef Full Text | Google Scholar
Knief,C. (2015). pmoAを分子マーカーとして評価した培養および未培養好気性メタン生成菌の多様性と生息地の嗜好性。 Front. Microbiol. 6:1346. doi: 10.3389/fmicb.2015.01346
PubMed Abstract | CrossRef Full Text | Google Scholar
Kotsyurbenko, O., Friedrich, M., Simankova, M., Nozhevnikova, A., Golyshin, P., Timmis, K.、他 (2007). 西シベリアの泥炭地における低 pH 値での酢酸依存性メタン生成から H2 依存性への移行と好酸性メタノバクテリウム菌株の単離。 Appl. Environ. Microbiol. 73, 2344-2348. doi: 10.1128/AEM.02413-06
PubMed Abstract | CrossRef Full Text | Google Scholar
Langenheder, S., and Székely, A. J. (2011). 細菌群集の初期形成には、種の選別と中立的なプロセスの両方が重要である。 ISME J. 5, 1086-1094. doi: 10.1038/ismej.2010.207
PubMed Abstract | CrossRef Full Text | Google Scholar
Liebner, S., Ganzert, L., Kiss, A., Yang, S., Wagner, D., and Svenning, M. M.(2015). “細菌群集の初期形成には、種の選別と中立的なプロセスが重要である。 不連続性永久凍土の劣化に伴うメタン生成コミュニティ組成とメタンフラックスの変化. Front. Microbiol. 6:356. doi: 10.3389/fmicb.2015.00356
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, Y. Q., Priscu, J. C., Xiong, J. B., Conrad, R., Vick-Majors, T., Chu, H. Y., et al(2016).。 Salinity drives archaeal distribution patterns in high altitude lake sediments on the Tibetan Plateau.(チベット高原の高地湖沼堆積物における塩分による古細菌の分布パターン)。 FEMS Microbiol. Ecol. 92:fiw033. doi: 10.1093/femsec/fiw033
PubMed Abstract | CrossRef Full Text | Google Scholar
Logares, R., Lindstrom, E. S., Langenheder, S., Logue, J. B., Paterson, H., Laybourn-Parry, J.、他(2013年). 進行性の長期的な環境変化にさらされた細菌群集の生物地理学。 ISME J. 7, 937-948. doi: 10.1038/ismej.2012.168
PubMed Abstract | CrossRef Full Text | Google Scholar
Loreau, M., Naeem, S., Inchausti, P., Bengtsson, J., Grime, J. P., Hector, A.、ほか(2001). 生物多様性と生態系機能:現在の知見と将来の課題。 Science 294, 804-808. doi: 10.1126/science.1064088
PubMed Abstract | CrossRef Full Text | Google Scholar
Lozupone, C. A., and Knight, R. (2007). 細菌の多様性におけるグローバルパターン。 Proc. Natl. Acad. Sci. U.S.A. 104, 11436-11440. doi: 10.1073/pnas.0611525104
PubMed Abstract | CrossRef Full Text | Google Scholar
Luton, P. E., Wayne, J. M., Sharp, R. J., and Riley, P. W. (2002). 埋立地のメタン菌集団の系統解析における16S rRNAに代わるmcrA遺伝子. Microbiology 148, 3521-3530. doi: 10.1099/00221287-148-11-3521
PubMed Abstract | CrossRef Full Text | Google Scholar
Lutz, S., Anesio, A. M., Raiswell, R., Edwards, A., Newton, R. J., Gill, F.、他 (2016).All. 紅雪マイクロバイオームの生物地理学と北極氷河の融解におけるその役割。 Nat. Commun. 7:11968. doi: 10.1038/ncomms11968
PubMed Abstract | CrossRef Full Text | Google Scholar
Martiny, J. B. H., Bohannan, B. J. M., Brown, J. H., Colwell, R. K., Fuhrman, J. A., Green, J. L.、他 (2006). 微生物生物地理学:微生物を地図上に表示する。 Nat. Rev. Microbiol. 4, 102-112. doi: 10.1038/Nrmicro1341
PubMed Abstract | CrossRef Full Text | Google Scholar
Martiny, J. B. H., Eisen, J. A., Penn, K., Allison, S. D. および Horner-Devine, M.C. (2011). バクテリアのベータ多様性の推進力は空間スケールに依存する。 Proc. Natl. Acad. Sci. U.S.A. 108, 7850-7854. doi: 10.1073/pnas.1016308108
PubMed Abstract | CrossRef Full Text | Google Scholar
McArdle, B. H., and Anderson, M.J. (2001). 地域社会データへの多変量モデルの当てはめ:距離に基づく冗長性分析についてのコメント。 Ecology 82, 290-297. doi: 10.1890/0012-9658(2001)0822.0.CO;2
CrossRef Full Text | Google Scholar
McCalley, C. K., Woodcroft, B. J., Hodgkins, S. B., Wehr, R. A., Kim, E.-H., Mondav, R., et al(2014).。 永久凍土融解に対する微生物群集の反応によって制御されるメタンダイナミクス。 Nature 514, 478-481. doi: 10.1038/nature13798
PubMed Abstract | CrossRef Full Text | Google Scholar
McGenity, T. J. (2010). “Methanogens and Methanogenesis in Hypersaline Environments,” in Handbook of Hydrocarbon and Lipid Microbiology, ed. K. N. Timmis (Berlin: Springer), 665-680. doi: 10.1007/978-3-540-77587-4_53
CrossRef Full Text | Google Scholar
McLusky, D. S. and Elliott, M. (2004). 河口域エコシステム. Ecology, Threats and Management(生態学、脅威と管理). New York, NY: doi: 10.1093/acprof:oso/9780198525080.001.0001
CrossRef Full Text | Google Scholar
Megonigal, J. P., Mines, M. E., and Visscher, P. T. (2005). 「嫌気性代謝:微量ガスと好気性プロセスとの関連性”, Biogeochemistry, ed. W. H. Schlesinger (Oxford: Elsevier), 350-362.
Google Scholar
Milferstedt, K., Youngblut, N. D., and Whitaker, R. J. (2010).「嫌気性代謝:微量ガスと好気性プロセスの関連」, ed. Biogeochemistry, ed. 腐植沼湖におけるメタン生成菌集団の空間構造と持続性. ISME J. 4, 764-776. doi: 10.1038/Ismej.2010.7
PubMed Abstract | CrossRef Full Text | Google Scholar
Mosier, A. C., and Francis, C. A. (2008). サンフランシスコ湾河口域におけるアンモニア酸化アーキアおよびバクテリアの相対的存在量と多様性。 Environ. Microbiol. 10, 3002-3016. doi: 10.1111/j.1462-2920.2008.01764.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Nazaries, L., Murrell, J. C., Millard, P. , Baggs, L.and Singh, B. K.(2013). メタン,微生物,モデル:将来予測に向けた土壌メタンサイクルの基礎的理解. Environ. Microbiol. 15, 2395-2417. doi: 10.1111/1462-2920.12149
PubMed Abstract | CrossRef Full Text | Google Scholar
Nelson, M. B., Martiny, A. C., and Martiny, J. B. H. (2016). 土壌中の微生物窒素循環形質のグローバルバイオジオグラフィー. Proc. Natl. Acad. Sci. U.S.A. 113, 8033-8040. doi: 10.1073/pnas.1601070113
PubMed Abstract | CrossRef Full Text | Google Scholar
Nemergut, D. R., Costello, E. K., Hamady, M., Lozupone, C., Jiang, L., Schmidt, S. K.、他 (2011). 細菌分類群の生物地理学におけるグローバルパターン。 Environ. Microbiol. 13, 135-144. doi: 10.1111/j.1462-2920.2010.02315.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Nobu, M.K., Narihiro, T., Kuroda, K., Mei, R. and Liu, W.T. (2016). Chasing the elusive Euryarchaeota class WSA2: genomes reveal a uniquely fastidious methyl-reducing methanogen. ISME J. 10, 2478-2487. doi: 10.1038/ismej.2016.33
PubMed Abstract | CrossRef Full Text | Google Scholar
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P.R., O’Har, R.B., et al. (2015).VEGA: Community Ecology Package(コミュニティエコロジーパッケージ)。 Rパッケージバージョン2.2.0. で入手可能。 https://CRAN.R-project.org/package=vegan
Google Scholar
Oloo, F., Valverde, A., Quiroga, M. V., Vikram, S., Cowan, D., and Mataloni, G.(2016).,。 生息地の不均質性と連結性が南米泥炭地の微生物群集を形成している。 Sci. Rep. 6:25712. doi: 10.1038/srep25712
PubMed Abstract | CrossRef Full Text | Google Scholar
Pattnaik, P., Mishra, S. R., Bharati, K., Mohanty, S. R., Sethunathan, N. and Adhya, T. K. (2000). 熱帯稲作土壌におけるメタン生成と関連微生物叢に対する塩分の影響。 Microbiol. また、このような環境下において、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」を実現するためには、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「環境負荷の低減」、「国内外での環境負荷低減」、「国際的な環境負荷低減」、「国際的な環境負荷低減」を実現する必要がある。 潮間帯湿地からのメタン排出に対する塩分の影響. Wetlands 31, 831-842. doi: 10.1007/s13157-011-0197-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Potter, E. G., Bebout, B. M., and Kelley, C. A. (2009).メタン排出の塩分濃度変化。 超過塩水微生物マット系における塩分勾配に沿ったメタンと推定されるメタン生成基質の同位体組成. Astrobiology 9, 383-390. doi: 10.1089/ast.2008.0260
PubMed Abstract | CrossRef Full Text | Google Scholar
R Core Team (2014). R: A Language and Environment for Statistical Computing (Version 3.0.3). Vienna: R Foundation for Statistical Computing.
Google Scholar
Roberts, D. (2016). labdsv: Ordination and Multivariate Analysis for Ecology(生態学のための順序付けと多変量解析)。 R Package Version 1.8-0.で入手可能。 https://CRAN.R-project.org/package=labdsv
Google Scholar
Rooney-Varga, J. N., Giewat, M. W., Duddleston, K. N., Chanton, J. P., and Hines, M. E. (2007)(2017. アラスカ泥炭地における古細菌群集構造、植生タイプ、メタン生成経路の関連性。 FEMS Microbiol. Ecol. 60, 240-251. doi: 10.1111/j.1574-6941.2007.00278.x
PubMed Abstract |Ref Full Text | Google Scholar
Rosenberg, E.DeLong, E.F., Lory, S.Stackebrandt, E. and Thompson, F. (2014). 原核生物の バクテリアの他の主要な系統とアーキア. ベルリン: Springer.
Google Scholar
Ruff, S. E., Biddle, J. F., Teske, A. P., Knittel, K., Boetius, A., and Ramette, A. (2015).原核生物:その他の主要系統と古細菌. メタン湧出微生物群のグローバルな分散とローカルな多様化。 Proc. Natl. Acad. Sci. U.S.A. 112, 4015-4020. doi: 10.1073/pnas.1421865112
PubMed Abstract | CrossRef Full Text | Google Scholar
Schauer, R., Bienhold, C., Ramette, A., and Harder, J. (2009). 南大西洋の深海表層堆積物におけるバクテリアの多様性と生物地理学的特徴. ISME J. 4, 159-170. doi: 10.1038/ismej.2009.106
PubMed Abstract | CrossRef Full Text | Google Scholar
Schloss, P. D., Westcott, S. L., Ryabin, T., Hall, J. R., Hartmann, M., Hollister, E. B., and al. (2009)。 このような場合、「萌芽的研究」は、「萌芽的研究」を「萌芽的研究」と呼ぶことにします。 Appl. Environ. Microbiol. 75, 7537-7541. doi: 10.1128/Aem.01541-09
PubMed Abstract | CrossRef Full Text | Google Scholar
Statham, P. J. (2012). 河口における栄養塩-概要と気候変動による潜在的影響。 Sci. Total Environ. 434, 213-227. doi: 10.1016/j.scitotenv.2011.09.088
PubMed Abstract | CrossRef Full Text | Google Scholar
Thauer, R. K., Kaster, A.-K. , Seedorf, H. , Buckel, W., and Hedderich, R. (2008). メタン生成古細菌:エネルギー保存における生態学的関連性の違い。 Nat. Rev. Microbiol. 6, 579-591. doi: 10.1038/Nrmicro1931
PubMed Abstract |Ref Full Text | Google Scholar
Tilman, D., Isbell, F., and Cowles, J.M. (2014). 生物多様性と生態系機能. Annu. Rev. Ecol. Evol.Syst. 45, 471-493. doi: 10.1146/annurev-ecolsys-120213-091917
CrossRef Full Text | Google Scholar
Ulrich, W., Hajdamowicz, I., Zalewski, M., Stañska, M., Ciurzycki, W., and Tykarski, P. (2010). 種混合か生息域のフィルタリングか:湖の島々のクモ群集のケーススタディ。 Ecol. Res. 25, 375-381. doi: 10.1007/s11284-009-0661-y
CrossRef Full Text | Google Scholar
Vanwonterghem, I., Evans, P.N., Parks, D.H., Jensen, P.D., Woodcroft, B.J., Hugenholtz, P., et al(2016).の項参照。 古細菌門Verstraetearchaeotaで発見されたメチロトローフ・メタン生成。 Nat. Microbiol. 1:16170. doi: 10.1038/nmicrobiol.2016.170
PubMed Abstract | CrossRef Full Text | Google Scholar
von Mering, C., Hugenholtz, P., Raes, J., Tringe, S. G. , Doerks, T., Jensen, L. J. , et al. 多様な環境における微生物群集の定量的な系統評価。 Science 315, 1126-1130. doi: 10.1126/science.1133420
PubMed Abstract | CrossRef Full Text | Google Scholar
Wagner, D., and Liebner, S. (2009). 「地球温暖化と永久凍土の炭素動態:メタン生成と酸化”, in Permafrost Soils, ed. R. Margesin (Berlin: Springer), 219-236.
Google Scholar
Weiher, E., and Keddy, P. (2001). エコロジカル・アセンブリー・ルール: Perspectives, Advances, Retreats. Cambridge: Google Scholar
Whitaker, R. J., Grogan, D. W., and Taylor, J. W. (2003). 地理的な障壁が超好熱性古細菌の固有個体群を隔離している。 Science 301, 976-978. doi: 10.1126/science.1086909
PubMed Abstract | CrossRef Full Text | Google Scholar
Wickham, H. (2009). ggplot2.NET。 Elegant Graphics for Data Analysis. New York, NY: Springer-Verlag. doi: 10.1007/978-0-387-98141-3
CrossRef Full Text | Google Scholar
Yang, J., Ma, L. A., Jiang, H., Wu, G., and Dong, H. (2016)(2016). 塩分は青海チベット湖の表層堆積物における微生物多様性と群集構造を形成する。 Sci. Rep. 6:25078. doi: 10.1038/srep25078
PubMed Abstract | CrossRef Full Text | Google Scholar
Yang, S., Liebner, S., Alawi, M., Ebenhöh, O., and Wagner, D. (2014). MOTHURによるmcrA遺伝子454パイロシークエンスデータ処理のための分類学データベースとカットオフ値。 J. Microbiol. Methods 103, 3-5. doi: 10.1016/j.mimet.2014.05.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Yang, S., Liebner, S., Winkel, M., Alawi, M., Horn, F., Dörfer, C., et al.(2017).を参照。 高標高の永久凍土の影響を受けた湿地から採取したコアメタン生成コミュニティの詳細な分析。 Soil Biol. Biochem. 111, 66-77. doi: 10.1016/j.soilbio.2017.03.007
CrossRef Full Text | Google Scholar
Ye, R. Z., Jin, Q. S., Bohannan, B., Keller, J. K., McAllister, S. A.., ピートランドにおける嫌気性炭素無機化,メタン生成効率,メタン生成経路に対するpH制御は,ombrotrophic-minelotrophic勾配を越えて行われた。 Soil Biol. Biochem. 54, 36-47. doi: 10.1016/j.soilbio.2012.05.015
CrossRef Full Text | Google Scholar
Zinder, S. H. (1993). “メタン生成菌の生理生態”, メタン生成: 生態学、生理学、生化学&遺伝学、ed.Methanogenesis: Ecology, Physiology, Biochemistry & Genetics. J. G. Ferry (Boston, MA: Springer), 128-206.
Google Scholar
Zinger, L., Amaral-Zettler, L. A., Fuhrman, J. A., Horner-Devine, M. C., Huse, S. M., Welch, D. B. M., et al. (2011). 海底・海水生態系におけるバクテリアのベータ多様性のグローバルパターン。 PLoS ONE 6:e24570. doi: 10.1371/journal.pone.0024570
PubMed Abstract | CrossRef Full Text | Google Scholar
.