Die meisten von uns haben in der Schule gelernt, dass es drei Arten lebender Säugetiere gibt – Eutertiere, Beuteltiere und Kloakentiere – und dass die offensichtlichsten Unterschiede zwischen ihnen darin bestehen, wie sie sich fortpflanzen. Die eutherischen oder „plazentaren“ Säugetiere, wie der Mensch, machen die große Mehrheit der heutigen Säugetiervielfalt aus. Alle Eutherier haben eine Chorioallanto-Plazenta, ein bemerkenswertes Organ, das sich nach der Empfängnis an der Stelle bildet, an der der Embryo mit der Gebärmutterschleimhaut der Mutter in Kontakt kommt (Langer, 2008).
Beuteltiere und Einhufer gehen mit der Schwangerschaft unterschiedlich um (Abbot und Rokas, 2017; Renfree, 2010). Eierlegende Monotremen, wie das Schnabeltier, haben winzige „Kugeln“, die aus ledrigen Schalen schlüpfen. Beuteltiere – Kängurus, Koalas, Bandicoots, Opossums usw. – haben Lebendgeburten, aber ihre Schwangerschaften sind kurz und ihre winzigen Jungen sind entwicklungsmäßig unreif und scheinen kaum Bedarf an einer Plazenta zu haben. Nach der Geburt entwickeln sich die Jungen außerhalb des Körpers ihrer Mutter weiter, oft in Falten und Beuteln am Bauch der Mutter.
Bei Beuteltieren ist die Milch, die die Mutter nach der Geburt gibt, von zentraler Bedeutung für die Entwicklung des Nachwuchses, und anders als bei eutherischen Säugetieren ändert sich die Zusammensetzung dieser Milch dramatisch, wenn die Jungen wachsen. Im Wesentlichen übernehmen die Milchdrüsen von Beuteltieren viele der Funktionen der Plazenta von Eutern (Renfree, 2010; Sharp et al., 2017; Abbildung 1). Und um das, was Sie vielleicht im Biologieunterricht gelernt haben, noch mehr auf den Kopf zu stellen: Beuteltiere haben zwar eine Plazenta, aber sie entwickelt sich erst spät in der Schwangerschaft und aus anderen Geweben als bei Euthertiern. Der Unterschied zwischen eutherischen Säugetieren und Beuteltieren besteht also nicht im Vorhandensein oder Nichtvorhandensein einer Plazenta, sondern in der relativen Bedeutung, die der Plazentation und der Laktation zukommt, um die Nachkommen durch die Entwicklung zu bringen.
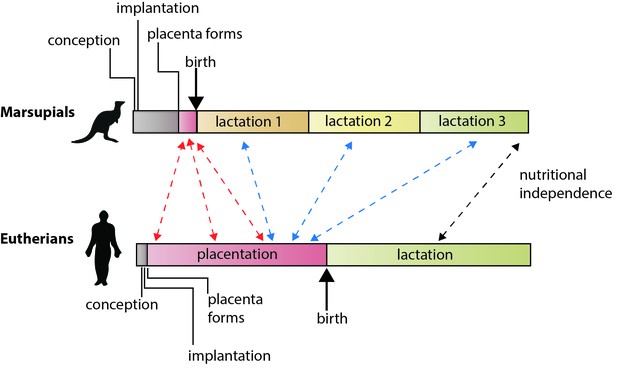
Die unterschiedlichen Fortpflanzungsstrategien von eutherischen Säugetieren und Beuteltieren.
Bei eutherischen Säugetieren ist die von der Mutter in die Aufzucht der Jungen vor der Geburt (über die Plazentation) und nach der Geburt (über die Laktation) investierte Energie ungefähr gleich groß. Bei Beuteltieren ist die Trächtigkeit kurz, die Plazenta bildet sich erst spät in der Schwangerschaft, und die Stillzeit ist länger. Guernsey et al. zeigen, dass die genetischen Merkmale, die die Entwicklung über die Plazenta bei Eutertieren regulieren, mit der kurzlebigen Beuteltierplazenta gemeinsam sind (rote Pfeile). Sie zeigen auch, dass einige der Gene, die den Plazentafunktionen bei Eutertieren zugrunde liegen, während der Laktation bei Beuteltieren exprimiert werden (blaue Pfeile), einschließlich verschiedener konservierter Komponenten der Laktation selbst (schwarzer Pfeil; Lefèvre et al., 2010). Hinweis: Die Zeitskalen sind nicht absolut.
Jetzt berichten Julie Baker von der Stanford University School of Medicine, Marilyn Renfree von der University of Melbourne und Mitarbeiter – darunter Michael Guernsey von Stanford als Erstautor, Edward Chuong von der University of Utah und Guillaume Cornelis (Stanford) – in eLife über neue Details der molekularen Mechanismen, die der Plazentation und Laktation bei Eutheriern und Beuteltieren zugrunde liegen (Guernsey et al., 2017). Die Ergebnisse wurden mit einer modifizierten Version einer Technik namens RNA-seq erzielt, um zu messen, wie sich das Transkriptom (der vollständige Satz von RNA-Transkripten in einer Zelle oder einem Satz von Zellen) zwischen verschiedenen Zelltypen während der Entwicklung verändert (Rokas und Abbot, 2009).
Guernsey et al. verglichen Veränderungen in der Genexpression in zwei Zelltypen in der Plazenta eines Tammar Wallaby, einem kleinen australischen Beuteltier, während der Entwicklung. Sie stellten fest, dass sich die Genexpression in den beiden Geweben unterschied und sich zudem im Laufe der Zeit dynamisch veränderte, ähnlich wie bei Eutertieren. Darüber hinaus fanden sie unter den Transkripten viele, die in der Plazenta von Euthern entscheidende Funktionen haben, darunter Mitglieder der Ig7-Signalwege und GCM1, ein Transkriptionsfaktor, der für die Bildung und Entwicklung der Plazenta wichtig ist. Und es waren nicht nur die Gene, die konserviert waren, die Muster der Genexpression in der Wallaby-Plazenta ähnelten denen, die in der Plazenta der Maus in den frühen Stadien der Schwangerschaft zu sehen sind. Dies ist genau das, was wir erwarten würden, wenn die Plazenta beim Wallaby frühe Entwicklungsfunktionen übernimmt und spätere Funktionen erst nach der Geburt erfüllt werden. Dies deutet darauf hin, dass ein wesentlicher Unterschied zwischen Beuteltieren und Eutertieren nicht in den frühen Funktionen der Plazenta liegt, sondern vielmehr darin, wie die Plazentafunktionen im Laufe der Evolution der Eutertierschwangerschaft aufgeteilt wurden.
Schließlich charakterisierten Guernsey et al. die Muster der Genexpression in den Brustdrüsen des Tammar und verschiedener Säugetiere. Sowohl die Maus als auch das Wallaby wiesen ähnliche Muster der Genexpression auf, was das Thema der funktionellen Kompartimentierung und Erhaltung in beiden Gruppen unterstreicht. Besonders bemerkenswert ist jedoch, dass die Forscher eine Reihe von Genen in den Brustdrüsen des Tammar identifizierten, von denen bekannt ist, dass sie eine wichtige Funktion in der Plazenta von Eutertieren haben (Abbildung 1). Zu diesen Genen gehören Gene, die am Nährstofftransport beteiligt sind, sowie mehrere Gene, von denen bekannt ist, dass sie für die Plazentation bei Euthern erforderlich sind (einschließlich GCM1). Diese Übereinstimmung der Genexpression deutet darauf hin, dass bei Beuteltieren die Plazenta die frühe fötale Entwicklung und die Laktation die späte fötale Entwicklung steuert, wobei einige der gleichen Gene und molekularen Pfade wie bei Eutern verwendet werden.
Diejenigen, die sich mit Beuteltieren befassen, haben seit langem argumentiert, dass wir unsere Lehrbücher korrigieren müssen, um die Plazenta der Beuteltiere und ihre ausgeprägt komplexe Laktation anzuerkennen (Renfree, 1983). Guernsey et al. untermauern dies, indem sie zeigen, dass sowohl Eutherier als auch Beuteltiere ein konserviertes Instrumentarium an Genen exprimieren, die zwar in unterschiedlichen Geweben und Organen lokalisiert sind, aber gemeinsamen Zwecken in der fötalen Entwicklung dienen. Diese überraschende Konservierung unterstreicht die Bedeutung der Identifizierung der Gene, die den funktionellen Veränderungen im Laufe der Evolution zugrunde liegen (Rausher und Delph, 2015).
Mit Blick auf die Zukunft ist es erwähnenswert, dass Beuteltiere in Bezug auf reproduktive Merkmale enorm variieren (Tyndale-Briscoe, 2005), und dass die Charakterisierung weiterer Arten in der Art und Weise, wie es Guernsey et al. für das Tammar-Wallaby getan haben, ein umfassenderes Verständnis der Evolution und Vielfalt der Beuteltierschwangerschaft selbst liefern wird. Es sind jedoch weitere Arbeiten erforderlich, um geeignete statistische Methoden zur Quantifizierung der Erhaltung von Transkriptom-Profilen zwischen den Arten zu entwickeln. Über die Säugetiere hinaus gibt es Formen der Plazentation bei Eidechsen, Seepferdchen und Insekten, und erste Studien deuten darauf hin, dass viele der beteiligten Gene oder Merkmale gemeinsam sind (Ostrovsky et al., 2016; Whittington et al., 2015). Es wird faszinierend sein zu erfahren, wie tief wir die Ursprünge des Schwangerschaftsbaukastens zurückverfolgen können.