Introduktion
Metan (CH4) är en viktig växthusgas. Utsläpp från naturliga miljöer som våtmarker, hav och sediment står för över 70 % av metanet i atmosfären globalt sett (IPCC, 2007). En bedömning av publicerade uppgifter visade att metanutsläppen varierar mellan våtmarker, sjöar, floder, flodmynningar och hav (i fallande ordning, kompletterande figur S1). Naturliga våtmarker står ensamma för 62 % av den biogena CH4-produktionen (Kirschke et al., 2013; Nazaries et al., 2013) och våtmarksutsläpp dominerar metankällornas mellanårsvariabilitet (Bousquet et al., 2006). Den stora arealen marina ekosystem bidrar däremot endast med cirka 8 % till de naturliga CH4-källorna (Nazaries et al., 2013). Av de globala oceaniska metanutsläppen bidrar flodmynningar med mindre än 10 % (Bange et al., 1994).
Metanogenes, den biologiska bildningen av metan, utförs av metanogena arkéer som producerar metan främst från H2/CO2, metylgrupper eller acetat vid anoxiska förhållanden (Thauer et al., 2008). Denna reaktion katalyseras av metylcoenzym M-reduktas (MCR). Genen mcrA som kodar för en underenhet av detta enzym är en vanligt förekommande genmarkör i molekylära undersökningar (Conrad, 2007; Bridgham et al., 2013). Fördelen med mcrA-genmarkören är att den fångar både de fylogenetiska och funktionella signaturerna hos metanogener, vilket ger ett stort sekvenseringsdjup för denna särskilda funktion (Luton et al., 2002; Borrel et al., 2013; Yang et al., 2014). Ett stort antal mcrA-sekvenser hämtades från olika naturliga miljöer. Den offentliga mcrA-databasen gör det möjligt att extrahera allmänna ekologiska mönster och undersöka de formande miljögradienterna på globala och regionala skalor. Dessutom finns en databas som sammanfattar de fysiologiska egenskaperna hos 152 metanogena isolat1 (Jabłoński et al., 2015). Nyligen avslöjade man genom binning ovanliga mcrA-sekvenser i den nya klassen Methanofastidiosa (Nobu et al., 2016) och de nya fylerna Bathyarchaeota (Evans et al., 2015) och Verstraetearchaeota (Vanwonterghem et al., 2016). Dessa nya fynd utökade vår kunskap om mångfalden av potentiella metanogener men skymde inte tillämpligheten av mcrA-genen som en molekylär markör för den stora majoriteten av metanogena samhällen.
Hittills har metanogena samhällen upptäckts i våtmarker, sediment, permafrostområden, risfält, rötkammar, rötkammaranläggningar, geotermiska källor och hydrotermiska skorstenar (Conrad, 2007; Thauer et al., 2008; Wagner och Liebner, 2009). Den metanogena samhällsstrukturen visade sig vara förknippad med miljöns pH, temperatur, salthalt, grundvattennivå och vegetationsdynamik på olika rumsliga och tidsmässiga skalor (Megonigal et al., 2005; Milferstedt et al., 2010; Frank-Fahle et al., 2014; McCalley et al., 2014; Cui et al., 2015; Liebner et al., 2015). Till exempel hindras acetoklastisk metanogenes i allmänhet av lågt pH eftersom det minskar acetatdissociationen (Megonigal et al., 2005; Kotsyurbenko et al., 2007). Vegetationen kan tillhandahålla labilt organiskt kol av hög kvalitet för att ge bränsle till metanogener i form av rotexudat eller detritus så att växtexudat i allmänhet gynnar acetoklastiska metanogener främst i myrar (Bridgham et al., 2013). Sulfat från havsvatten hämmar metanproduktionen i våtmarker med tidvatten, och salthalten har därför använts som en allmän prediktor för metanutsläpp (Holm et al., 2016). En studie på tibetanska sjösediment visade att ökad salthalt hämmar hydrogenotrofa metanogener men ökar den acetoklastiska metanogenesen (Liu et al., 2016). Dessa studier visade på miljömässiga drivkrafter för metanogena samhällen, men har fokuserat på enstaka livsmiljöer eller begränsade rumsliga skalor.
För att förstå metanogenernas anpassning till olika miljöförändringar krävs dock en systematisk och global utforskning av korrelationerna mellan sammansättningen av mikrobiella samhällen och miljöförhållanden (Lozupone och Knight, 2007). För närvarande är det endast ett fåtal studier som behandlar spridning och habitatfiltrering av metanogena samhällen (Auguet et al., 2010; Barreto et al., 2014). Vi antar att metanogena sammansättningar huvudsakligen påverkas av habitatfiltrering och att detta styrs av globala miljökontroller. Med tanke på att metanutsläppshastigheterna skiljer sig mycket åt mellan naturliga ekosystem kan en explicit integrering av sammansättningen, mångfalden och biogeografin hos metanogena sammansättningar i dessa ekosystem vara grundläggande för att bestämma metanproduktionens svar på nuvarande och framtida klimatförändringar. Denna metastudie utförs för att fylla de luckor som är förknippade med metanogen biogeografi, mångfald och dess miljökontroller genom att använda offentligt tillgängliga mcrA-sekvensdata och litteratur som kompletteras med fysiologiska data från metanogena isolat.
Material och metoder
Datainsamling
Vi hämtade mcrA-sekvenser som var tillgängliga i GenBank (januari 2016)2. För varje träff kontrollerades originalhandlingen och motsvarande mcrA-sekvenser parsades av ett anpassat Perl-skript. Eftersom vi fokuserade på naturliga miljöer erhölls metanogena mcrA-sekvenser från naturliga livsmiljöer och klassificerades som jord, sjö-, flodmynning-, havs- och hydrotermala sediment samt lervulkaner. Fem bibliotek från nästa generations sekvensering (NGS) inkluderades utöver sekvenser från klonbibliotek. Sekvenserna hämtades utan att ta hänsyn till den relativa abundansen i det ursprungliga datasetet. Eftersom sekvenser från klonbibliotek huvudsakligen täcker de rikliga fylotyperna medan NGS kan fånga en mycket djupare mångfald, gjorde vi en kompromiss för att använda NGS-data men mildrade ett potentiellt fel på grund av sekvenseringsmetodernas olika upplösning. Därför valde vi endast representativa sekvenser för rikligt förekommande OTU:er med en relativ förekomst på mer än 1 %. Vi avvisade också de NGS-sekvenser som inte klarade översättningskontrollen från nukleotid- till proteinsekvenser eller som hade låg kvalitet (sekvenser < 250 bps). Slutligen konstruerade vi ett dataset som innehöll 4466 unika mcrA-sekvenser från 94 globalt spridda platser (figur 1 och kompletterande tabeller S1, S2). Dessutom subtraherade vi inte mcrA-sekvenser från potentiella arkealiska metanotrofer från datasetet, vilket oundvikligen upptäcktes i den genomiska undersökningen (Conrad, 2007). Denna del ligger utanför den här studiens fokus.
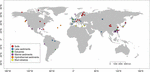
FIGUR 1. Placering av de platser som använts i denna undersökning. Platserna grupperades i sex olika kategorier beroende på deras livsmiljöer och märktes med olika färger. En sammanfattning av de sex livsmiljöerna finns i de kompletterande tabellerna S1, S2.
De geografiska koordinaterna och miljöinställningarna, inklusive pH, salthalt, höjd, genomsnittlig årlig lufttemperatur (MAAT) och genomsnittlig årlig nederbörd (MAP), hämtades för varje undersökningsområde som beaktas i den här studien från motsvarande publikationer, förutsatt att uppgifterna är tillgängliga (se kompletterande tabell S1). För att ta hänsyn till saknade miljöparametrar i flera studier definierade vi kvalitativt vissa miljövariabler enligt platsbeskrivningarna i den relevanta litteraturen och konverterade sedan dessa kategoridata till semimetriska numeriska värden, t.ex. definierade vi marina sediment, hydrotermala sediment, vulkanisk lera och sedimentprover från sodasjöar som ”salthaltiga”, jord- och sedimentprover från sötvattensjöar som ”icke-salthaltiga”, och prover från mangrove- och flodmynningar som ”blandade” prover.
Råsekvensbearbetning
Sekvensbearbetningen genomfördes med mjukvaruplattformen Mothur (Schloss et al, 2009). Sekvenser från olika bibliotek sammanfördes före bearbetningen. Sekvenser med en längd på mindre än 350 bp eller mer än 8 tvetydiga baser kasserades. Därefter anpassades dessa sekvenser mot en förinställd delmängd av mcrA-sekvenser som hämtades från FunGene-databasen på http://fungene.cme.msu.edu/ (Fish et al., 2013). Chimeriska sekvenser identifierades med programvaran Mothur med hjälp av uchime-metoden (Edgar et al., 2011) med själva datasetet som referens. Därefter användes de giltiga nukleotidsekvenserna för mcrA-genen för att beräkna okorrigerade parvisa avstånd mellan de anpassade DNA-sekvenserna och tilldelades vidare till operativa taxonomiska enheter (OTU) med ett gränsvärde på 84 %, vilket motsvarar 97 % för 16S rRNA-genen (Yang et al., 2014). Förekomsten av varje mcrA OTU redovisades endast som närvaro eller frånvaro. Vi ökade noggrannheten i den taxonomiska klassificeringen av OTU:erna genom att ta hänsyn till både nukleotid- och aminosyrasekvenser. På DNA-nivå tilldelades den taxonomiska identiteten av Mothur-plattformen enligt en referensdatabas (Yang et al., 2014). På proteinnivå användes de anpassade proteinsekvenserna för att konstruera ett träd i ARB, och sedan baserades den taxonomiska tilldelningen på motsvarande databas. Om tilldelningen av en OTU var inkonsekvent, blåste vi manuellt både nukleotid- och proteinsekvensen i NCBI och fastställde den slutliga taxonomiska identiteten genom att ta hänsyn till frågetäckningen (>95 %), identiteten (>84 %) och e-värdet (<1E-5). För proteinsekvenser hänvisade avgränsningen på genusnivå till tröskelvärdet 83,5 % (Hunger et al., 2011).
Ökologisk och statistisk analys
Den statistiska analysen gjordes med hjälp av olika R-paket. Principal coordinates analysis (PCoA) ordineringar genererades baserat på Jaccard distansmatriser som konstruerades med hjälp av vegan-paketet v2.2.0 (Oksanen et al., 2015). Permutationell MANOVA (multivariat variansanalys) genomfördes för att bedöma källan till variation i Jaccardmatrisen (McArdle och Anderson, 2001) i vegan med 104 permutationer. Jaccardavståndsmåtten baseras på artens närvaro/frånvaro, vilket är mer lämpligt för vårt dataset eftersom de flesta studier endast tillhandahöll de representativa sekvenserna medan information om abundans saknas. De taxonomiska förekomstfrekvenserna över olika livsmiljöer visualiserades genom bubbelplottar med ggplot2-paketet (v1.0.0) (Wickham, 2009). Hierarkisk klusteranalys av de icke-saliska jord- och sjösedimentsamhällena utfördes med R-funktionen ”hclust” (R Core Team, 2014). De erhållna samhällsklusterna beskrevs enligt pH- och temperaturregimen i de ursprungliga proverna eftersom vi med hjälp av PCoA i förväg identifierade att båda parametrarna pH och temperatur påverkar sammansättningen av metanogena samhällen i icke-salina jordar och sjösediment. Varje metanogena linjens koppling till vart och ett av dessa kluster fastställdes med hjälp av korrelationsbaserad indikatorartsanalys (Dufrene och Legendre, 1997). Indikatorarter definieras här som arter som både förekommer rikligt i en viss typ av livsmiljö (specificitet) och som huvudsakligen återfinns i denna typ av livsmiljö (trohet). I den här studien valdes indikatortaxa, i likhet med indikatorartsbegreppet, för icke-saliska jordar och sjösediment ut i enlighet med ett indikatorvärde (IndVal-värde) med hjälp av R-paketet labdsv (Roberts, 2016) om sannolikheten för att få ett indikatorvärde som är lika högt som det som observerats under de angivna iterationerna är mindre än 0,05. Chao2-indexen beräknades för varje prov med hjälp av paketet vegan. Wilcoxon rangsummetestet av Chao2-index mellan livsmiljöer utfördes med R-funktionen ”wilcox.test” (R Core Team, 2014). För att befästa habitatfiltreringens inverkan på isolat av metanogena arkéer hämtades de fysiologiska och biokemiska egenskaperna hos beskrivna metanogena kulturer från3 ”Methanogenic archaea database” (Jabłoński et al., 2015). Bland dem filtrerades, kategoriserades och plottades isolaten med information om optimalt NaCl-behov enligt deras isoleringskälla.
För att undersöka spridningsbegränsningens påverkan på den metanogena samhällsstrukturen utfördes en linjär regressionsanalys baserad på en matris för geografiskt avstånd och en matris för Jaccard-avstånd mellan samhällen med hjälp av R-funktionen ”lm” (R Core Team, 2014). Vi utförde Mantel- och partiella Mantel-tester för att utvärdera effekterna av spridningsbegränsning enligt de två matriserna igen med hjälp av vegan-paketet i R (Oksanen et al., 2015). Vidare tillämpades multivariat rumslig analys (spatial PCA) på 16 europeiska jord- och sjösedimentprover baserat på Morans I-index för att utforska den rumsliga strukturen hos metanogenerna med hjälp av funktionen ”multispati” i ade4-paketet i R (Dray och Dufour, 2007). Dessutom genomfördes Ward’s Minimum variance clustering som baserades på Jaccard distansmatrisen på dessa 16 prover med hjälp av R-funktionen ”hclust” (R Core Team, 2014) och vi projicerade vidare klusterresultaten på en geografisk karta. Den europeiska shapefilen för kartläggning på delstatsnivå finns tillgänglig i GSHHG-databasen (v2.3.6)4. Kartan genererades med hjälp av QGIS v2.18.25.
Resultat
Biogeografi av metanogena arkéer i naturliga miljöer
McrA-gensekvenserna från 94 globalt spridda naturliga miljöer hämtades. Platsen och ekosystemtypen för var och en av dessa 94 platser visas i figur 1. Incidensfrekvenserna (närvaro/frånvaro) för metanogena linjer slogs samman enligt ekosystemtyp och illustreras i figur 2. I korthet är Methanoregula det mest frekventa taxonet i jord, tillsammans med Methanobacterium, Methanosaeta, Methanocella, Methanomassiliicoccus och Methanosarcina. I sediment från flodmynningar upptäcktes ofta sekvenser från Methanosaeta, Methanobacterium, Methanoregula och Methanoculleus. I sjösediment förekom dessutom främst Methanoregula och Methanosaeta. I marina sediment är Methanoculleus och Methanosaeta de vanligaste linjerna, följt av Methanolinea.
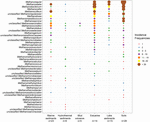
FIGUR 2. Bubbelplot som visar incidensfrekvenserna för metanogena linjer i olika naturliga miljöer. Rangordningen längs den vertikala axeln motsvarar de minskande totala incidensfrekvenserna för linjeformerna. Taxonomin visas på släktnivå. Om det inte var möjligt att hänföra till släktnivå användes den närmast högre taxonomiska nivån. Antalet prover (n) anges för varje livsmiljö.
Även om många taxa upptäcktes i olika miljöer visar vissa fortfarande miljöpreferenser. Methanoregula, det taxon som ofta förekommer i icke-marina miljöer och övergångsmiljöer (jord, sjösediment och flodmynningar), saknas i marina livsmiljöer (marina sediment, hydrotermiska sediment och lervulkaner). Methanobacterium och Methanocella, som förekommer i icke-marina miljöer och övergångsmiljöer, finns sällan i marina miljöer. Methanococcoides, som är en dominerande släktlinje i marina sediment, hydrotermiska sediment och lervulkaner, observeras däremot knappt i jordar och sjösediment. Methanogenium och Methanolacinia observeras dessutom endast i flodmynningar och marina sediment, medan Methanospirillum och Methanosphaerula endast förekommer i terrestra miljöer. Dessutom finns vissa specifika taxa uteslutande i hydrotermiska sediment, däribland Methanocaldococcus, Methanothermococcus, Methanopyrus, Methanotorris och Methanococcus. Även om vissa släktskap, t.ex. Methanosaeta, förekommer i de flesta miljöer kan inget släktskap anses vara allestädes närvarande.
Den största rikedomen av släktskap förekom i sediment från flodmynningar som också hyser jämnare incidensfrekvenser av olika släktskap. Däremot uppvisar lervulkaner och hydrotermiska ekosystem en relativt låg metanogen mångfald. Jordar och sjösediment som liknar flodmynningar kännetecknas av olika metanogena sammansättningar.
Alpha-diversitet hos metanogena samhällen i naturliga miljöer
Rikedomen av metanogena arkéer enligt Chao2-indexet varierade i hög grad mellan olika ekosystemtyper (figur 3). I syfte att direkt jämföra alfa-diversiteten och för att få en rimlig avvägning mellan proverna genomfördes en subsampling till 15 sekvenser för varje plats. Chao2-indexet visar att sediment i flodmynningar omfattar den högsta artrikedomen av metanogener bland de sex ekosystemtyperna (kompletterande tabell S3), vilket understryker resultaten från bubbeldiagrammet (figur 2). Jord- och sjösediment, som visar lägre rikedom än estuariaprover, har betydligt högre Chao2-index än marina sediment och hydrotermiska sediment och utan några signifikanta skillnader mellan marina sediment och hydrotermiska sediment (kompletterande tabell S3).
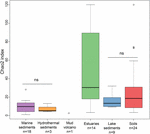
FIGUR 3. Boxplot av Chao2-index för de olika ekosystemtyperna. Plotten är baserad på delprovtagna dataset som innehåller 15 sekvenser för varje plats för att göra jämförelsen på alfa-diversitetsmått mer robust. Antalet prover i varje livsmiljö anges som ”n” under livsmiljöbeteckningen. ”ns” anger ingen statistisk signifikans i Wilcoxon-testet. Det statistiska resultatet av alfa-diversitet på OTU-nivå finns i kompletterande tabell S3.
Globala kontroller av metanogena samhällen i naturliga miljöer
De 94 globalt fördelade metanogena samhällena klustrades i en ordinationsdiagram genom att tillämpa PCoA baserat på Jaccard distansmatris. Enligt PCoA-analysen förklarar den första och andra axeln tillsammans 16,3 % av den totala variansen. Variationerna mellan proverna kan därmed till stor del förklaras av salthalt (figur 4). Eftersom det i vissa fall inte fanns några initiala uppgifter om saltkoncentrationerna, har vi kvalitativt klassificerat dessa prover som salthaltiga, blandade (intermediära) och icke-salthaltiga prover enligt beskrivningen ovan. De salthaltiga och icke-salthaltiga proverna skiljs effektivt åt längs den första axeln. De blandade proverna grupperar sig mellan de salthaltiga och icke-salthaltiga proverna. Den permutationella MANOVA som baseras på Jaccard distansmatrisen tyder också på att salthalt är den primära abiotiska faktorn som styr fördelningen av globala metanogena samhällen (R2 = 0,099, P < 0,001) (tabell 1).
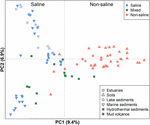
FIGUR 4. Principal coordinates analysis (PCoA) ordination baserad på Jaccard distansmatrisen för metanogena mcrA-gensekvensbibliotek där 94 prover jämförs. PCoA är färgad efter salthalt: De röda symbolerna indikerar icke-saltiga miljöer, de blå symbolerna indikerar salthaltiga miljöer och de gröna symbolerna indikerar intermediära miljöer. Olika symboler visar olika miljöer. Procentandelen av variationen som förklaras av de plottade huvudkoordinaterna anges på axlarna.

TABELL 1. Permutationell MANOVA-analys på Jaccard-distansmatrisen för alla prover från de sex livsmiljöerna för att testa sambandet mellan samhällets varians och olika miljövariabler.
Därtill kontrollerade vi om det fanns ett potentiellt samband mellan isoleringskällan för metanogena renodlade kulturer och den optimala koncentrationen av NaCl för tillväxt. Den optimala NaCl-koncentrationen för de metanogena renkulturerna uppvisade en minskning från marina till flodmynningar, sjösediment och markekosystem (kompletterande figur S2). Ett fåtal isolat med utfall kommer från sediment från sodasjöar eller hypersalina jordar.
Miljökontroller och metanogena indikatortaxa i icke-salina jordar och sjösediment
På global nivå är metanogena samhällen från icke-salina jordar och sjösediment tätt sammanfogade (figur 4), så vi analyserade ytterligare miljökontrollerna av metanogena samhällen från dessa två livsmiljöer, som totalt utgör 33 undersökningsområden. Gemenskapsbaserad klusteranalys för dessa två typer av livsmiljöer avslöjade fyra kluster baserade på Jaccard distansmatrisen (kompletterande figur S3). Permutationell MANOVA tyder på att både pH (R2 = 0,099, P < 0,001) och temperatur (R2 = 0,069, P < 0,001) påverkar den metanogena β-diversiteten i icke-saliska jordar och sjösediment (tabell 2). Följaktligen tilldelade vi de fyra klustren pH och MAAT på den ursprungliga provtagningsplatsen och fick i stort sett konsekventa undergrupper till samhällets klusterbildning (figurerna 5C, D). Kombinationen av miljöegenskaper och dessa fyra samhällskluster gör att vi kan definiera dessa fyra undergrupper som neutral och kall grupp 1, sur och kall grupp 2, sur och måttlig grupp 3 och neutral och varm grupp 4 (figur 5). Ytterligare PCoA-ordination baserad på Jaccard-dissimilaritetsmatrisen visar att längs PC1-axeln kommer de flesta proverna från grupp 2 och grupp 3 från sura jordar och sjösediment, medan grupp 1 och grupp 4 huvudsakligen kommer från neutrala miljöer (figur 5A). Dessutom skiljde sig prover från måttliga platser (grupp 3) från prover från varma och kalla platser längs PC2, medan proverna från varma miljöer (grupp 4) skiljde sig från de andra proverna längs PC3 (figur 5B). Därmed förklarar de tre första axlarna i PCoA-ordinationen 38,8 % av den totala variationen.

TABELL 2. Permutationell MANOVA baserad på en Jaccard-distansmatris för prover av icke-salint jord- och sjösediment för att testa sambandet mellan samhällets varians och olika miljövariabler.
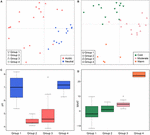
FIGUR 5. Plott för huvudkoordinatanalys baserad på en Jaccard-distansmatris för 33 icke-saliska jordar och sjösediment. Subplot (A) visar PC1 och PC2 och symbolerna är färgade efter pH, och subplot (B) visar PC2 och PC3 och symbolerna är färgade efter temperatur. De tre första komponenterna förklarar 17,1, 11,7 och 10 % av variansen. Boxplottarna visar pH (C) och MAAT (genomsnittlig årlig lufttemperatur) (D) för fyra identifierade undergrupper. Boxens färg i figur C motsvarar pH-kategorin i (A). Färgerna i boxplottarna visar statistisk signifikans baserat på ett parvis Wilcoxon-test (P < 0,05), där proverna med samma färg inte skiljer sig signifikant från varandra. På samma sätt följer färgen i (D) temperaturgrupperingen i (B). Undergrupperna hänvisar till den hierarkiska klusteranalysen av samhällets likheter.
Vi undersökte förekomsten av metanogena linjer i varje undergrupp baserat på incidensfrekvenser (figur 6). Methanoregula dominerar i alla typer av icke-salina livsmiljöer. Förutom Methanoregula uppvisar den neutrala och kalla undergruppen (grupp 1) en hög förekomst av Methanosaeta, Methanobacterium och Methanosarcina. Den sura och kalla gruppen 2 representeras av Methanobacterium, Methanocella och Methanosarcina medan Methanosaeta saknas här. Methanocella och Methanosaeta är vanliga i den sura och måttliga gruppen 3. I den neutrala och varma gruppen 4 är Methanolinea och Methanosaeta viktiga medlemmar. Denna grupp är den enda grupp där Methanoculleus identifierades.

FIGUR 6. Incidensfrekvenser för metanogena linjer inom de fyra definierade grupperna av metanogena samhällen i jord och sjösediment. Den vertikala axeln är ordnad i alfabetisk ordning. Staplarnas längd motsvarar de genomsnittliga incidensfrekvenserna för varje linje inom motsvarande grupp. Felstaplarna representerar standardavvikelsen för ett visst taxon över olika prover i den gruppen. Asterisker visar specialiserade taxa med P-värde < 0,05. Taxonomin visas för släktnivå. Om en tilldelning till släktnivå inte var möjlig användes nästa högre tilldelningsbara taxonomiska nivå. Beskrivningen för de fyra grupperna ges i figur 5.
För alla fyra grupperna är de taxa som har hög förekomst Methanoregula, Methanobacterium, Methanosarcina, Methanosaeta, Methanomassiliicoccus och Methanocella. De specialiserade taxa, som är betydligt mer representerade på de flesta av platserna inom en viss grupp, upptäcktes enligt analysen av indikatorarter enligt beskrivningen ovan. Totalt uppvisade sex av de 31 taxa ett signifikant indikatorvärde (P < 0,05) (markerade med asterisk i figur 6). Grupp 1 (neutral och kall) uppvisade det största antalet specialister med linjetyper av Methanosaeta, Methanolobus och Methanomethylovorans. Methanobacterium fungerade som specialisttaxon i den kalla och sura gruppen 2 medan Methanolinea identifierades som specialist i grupp 4 (neutral och varm) men observerades knappt i andra grupper. Dessutom är Methanoregula till stor del representerad i den sura och måttliga gruppen 3.
Dispersal Limitation
En linjär regressionsanalys (R2= 0,05, P < 0,001) visade på ett svagt samband mellan geografiskt avstånd och den metanogena samhällsstrukturen i det globala datasetet. Samtidigt visade ett Mantel-test att miljövariablerna har en högre korrelation med samhällsstrukturen än de geografiska avstånden (se tabell 3). Denna trend bekräftas också av ett partiellt Manteltest som kontrollerar autokorrelationseffekter. En plottning av det geografiska avståndet mot Jaccard-samhällets likhet visar ingen tydlig linjär trend utan mönster som huvudsakligen beror på den globala fördelningen av provtagningspunkterna (se kompletterande figur S4).

TABELL 3. Mantel- och partiella Manteltestanalyser för bestämning av inverkan av miljövariabler och geografiskt avstånd på den mikrobiella fördelningen för det globala datasetet och ett delprov av 16 europeiska prover.
För att ytterligare analysera påverkan av spridning begränsade vi vår analys till Europa, där proverna var tätast och mest jämna. Manteltest och partiella Manteltest på denna delmängd reproducerade trenden att samhällsdata är högre korrelerade till miljövariablerna än till geografiska avstånd (se tabell 3). Det partiella Manteltestet som kontrollerade miljövariablerna kunde inte påvisa någon statistiskt signifikant korrelation mellan det mikrobiella samhället och det geografiska avståndet. En rumslig PCA-analys av dessa 16 europeiska platser visar att det metanogena samhället har en rumslig struktur (23,7 % av den totala variansen förklarades av denna struktur), vilket motsvarar en positiv rumslig autokorrelation mellan platserna enligt Morans I-index (Morans I = 0,4018). Endast det första egenvärdet var stabilt och motsvarade en separation av proverna mellan Centraleuropa och de baltiska staterna (kompletterande figur S5). Den lilla datamängden försvårar dock en stabil tilldelning av denna observerade rumsliga struktur till geografiska variabler, miljövariabler eller till båda. Vi utförde därför en klusteranalys av de metanogena samhällena enligt beskrivningen ovan och avslöjade tre grupper (figur 7A) som vi projicerade på en geografisk karta (figur 7B). Klusterbildningen reproducerade inte uppdelningen av den rumsliga PCA:n längs Östersjön. Följaktligen uppvisar vissa platser som geografiskt sett ligger mycket nära varandra olika metanogena samhällsstrukturer och samlas i olika kluster. Å andra sidan uppvisar vissa geografiskt mycket avlägsna platser mycket likartade samhällssammansättningar och grupperar sig tillsammans (figur 7B). Den regionala spridningen av grupperna i Europa verkar inte motsvara eller begränsas av en geografisk struktur.
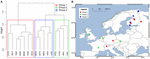
FIGUR 7. Hierarkisk klusteranalys av Jaccard distansmatrisen bland 16 europeiska jord- och sjösedimentprover. Ward-klustermetoden användes för analysen. Platserna delades in i tre grupper som färgades av olika rektanglar i klusterdendrogrammet (A) och projicerades på en europeisk karta (B).
Diskussion
Identifiering och tillämpning av biogeografiska begrepp på mikrobiella samhällen är av stort intresse inom mikrobiell ekologi. Mikrobiell biogeografi tros styras av det evolutionära och ekologiska samspelet mellan fyra huvudprocesser: habitatfiltrering, spridning, drift och mutation (Hanson et al., 2012). Även om inflytandet av drift och mutation ligger utanför den här studiens tolkningsförmåga visar vi att det finns globala mönster för metanogena samhällen i naturliga miljöer. Denna studie visar på ett globalt biogeografiskt mönster för metanogena samhällen som är mer förknippat med habitatfiltrering än med geografisk spridning. Metanogena samhällen från sediment från sodasjöar, till exempel, är nära sammankopplade med geografiskt avlägsna marina prover (figur 4) och mycket likartade metanogena samhällen förekommer i europeiska jord- och sjösediment trots att de är belägna på stort avstånd från varandra (figur 7). Sammantaget visade våra försök att avslöja potentiella spridningsbegränsningar att det geografiska läget har ett svagt inflytande på den metanogena samhällsstrukturen, vilket står i kontrast till ett tydligt inflytande från miljöförhållandena. Det är inte möjligt att slutgiltigt utesluta rumsliga effekter på de mikrobiella samhällena med de tillgängliga datapunkterna. Provtagningspunkterna är inriktade på vissa livsmiljöer och/eller områden medan det totala antalet prover är lågt. Om det finns en effekt antar vi att rumsliga effekter endast förekommer på regional eller lokal nivå. Lokala spridningsbegränsningar har rapporterats för hypertermofila arkéer på grund av geografiska hinder (Whitaker et al., 2003), ammoniakoxiderande bakterier i lokala saltsjöar, men inte på regional och kontinental skala (Martiny et al., 2011), mikroorganismer i djuphavssediment tillsammans med miljöförhållanden (Schauer et al., 2009), och för bakterier i en stor uppsättning heterogena snömiljöer som främst beror på tillgången på alloktoniskt kol (Lutz et al., 2016). Ett mindre inflytande av spridningsbegränsning på metanogener i naturliga miljöer innebär att metanogener kan fördela sig slumpmässigt i rummet, väljas framgångsrikt av den lokala miljön om deras fysiologiska krav uppfylls och kan etablera stabila samhällen (Martiny et al., 2006; von Mering et al., 2007).
Begreppet habitatfiltrering innebär att arter med liknande ekologiska krav bör förekomma tillsammans oftare än vad som kan förväntas av slumpen (Weiher och Keddy, 2001; Cornwell et al., 2006; Ulrich et al., 2010). Vårt resultat visar att det finns stora skillnader i sammansättningen av metanogena samhällen mellan marina ekosystem och ekosystem i våtmarker och sjöar, medan flodmynningar ligger däremellan. Biogeografiska mönster baserade på en uppsättning genundersökningar på miljöprover har också rapporterats för allmänna bakterier (Nemergut et al., 2011), mikrobiomet i metanläckor (Ruff et al., 2015), ammoniakoxiderande arkéer (Cao et al., 2013), marina pelagiska och bentiska bakterier (Zinger et al., 2011) och kvävecyklande mikroorganismer (Church et al., 2008). Habitatfiltrering har särskilt rapporterats för okultiverade arkéer (Auguet et al., 2010), hela bakteriesamhällen i olika miljöer (von Mering et al., 2007; Chaffron et al., 2010) eller i sydamerikanska torvmarker på regional skala (Oloo et al, 2016), liksom specifika bakteriegrupper som metanoxiderande (Knief, 2015) och kvävefixerande bakterier (Nelson et al., 2016).
Våra resultat visar att på global skala reglerar salthalten i hög grad det metanogena samhällets sammansättning och bestämmer stora skillnader mellan marina och terrestra metanogena sammansättningar. Även metanogener från sediment från sodasjöar grupperar sig med dem från marina sediment (figur 4), vilket belyser det globala inflytandet av salthalt. Detta resultat stämmer överens med andra studier baserade på 16S rRNA-genen som avslöjar att salthalt är en primär faktor som formar globala mönster av övergripande bakterie- och arkealsamhällen (Lozupone och Knight, 2007; Auguet et al., 2010; Caporaso et al., 2011; Cao et al., 2013). Ett lågt inflytande av geografisk separation men en stark inverkan av salthalt på allmänna mikrobiella samhällen observerades också i tidigare studier (Logares et al., 2013; Yang et al., 2016). Följaktligen avgör salthalten i stor utsträckning vilka linearier som kan överleva. I olika livsmiljöer var metanproduktionsaktiviteten negativt korrelerad med salthalten (Bartlett et al., 1987; Potter et al., 2009; Poffenbarger et al., 2011). Inhiberingen av metanproduktionen genom salthalt föreslås därmed sammanfalla med en minskad metanogen populationsstorlek (Pattnaik et al., 2000). Effekten av salthalt på hydrogenotrofisk, acetotrofisk och metylotrofisk metanogenes beror därmed på salthaltsnivån och är olika för de olika metanogenesens vägar (Liu et al., 2016). För närvarande finns det ingen tydlig mekanism för att förklara salthalts inverkan på samhällsstrukturen, men flera hypoteser kan fungera som möjliga förklaringar. Fysiologiskt sett påverkar salthalt cellernas externa och interna osmolaritet. De metanogena cellerna som inte är salthaltiga har utvecklat fysiologiska anpassningar för att motverka det inre turgatrycket, medan de saltanpassade cellerna har förlorat denna funktion (Zinder, 1993). Dessutom kan ökad salthalt få metanogenerna att syntetisera eller ta upp en ökad andel kompatibla lösningsmedel till en betydande energi- och därmed metabolisk kostnad (McGenity, 2010). Egenskapen salttolerans manifesteras till och med i den optimala NaCl-koncentrationen för tillväxt av metanogena renkulturer eftersom vi fann att isolaten från marina sediment och hydrotermiska sediment har betydligt högre optimal NaCl-koncentration än isolaten från jordar (Supplementary Figure S2).
I de icke-saltiga terrestra ekosystemen, särskilt i jordar och sjösediment, styrs det metanogena samhällets sammansättning av kombinationen av temperatur och pH. Följaktligen kan metanogenerna i dessa miljöer klassificeras i fyra grupper (figur 6). Till skillnad från marina ekosystem uppvisar de icke-salina terrestra ekosystemen en stor naturlig variation av både pH och temperatur. Temperaturen kan påverka inte bara den metanogena vägen utan även metanogena populationer i sig (Conrad, 2007; Rooney-Varga et al., 2007). Metanproduktionen kan öka kraftigt om temperaturen stiger till följd av de temperaturkänsliga stegen under fermentation och acetogenes (Megonigal et al., 2005; Kotsyurbenko et al., 2007). Dessutom kan ett lågt pH väsentligt begränsa tillgången på acetat genom att förhindra att acetat dissocieras och därmed påverka den acetoklastiska metanogenesen negativt (Fukuzaki et al., 1990; Bridgham et al., 2013). Detta kan vara en möjlig orsak till att Methanosaeta saknades i grupp2, medan Methanosarcina kan växla mellan olika källor och inte påverkades nämnvärt. Dessutom kan pH reglera effektiviteten av metanproduktion och metanogena vägar från ombrotrofa till minerotrofa torvmarker, genom direkt hämning av båda metanogenesvägarna och indirekt genom dess effekter på fermentationen (Ye et al., 2012). Därför kan både temperatur och pH direkt eller indirekt reglera de metaboliska stegen i samband med metanogenesen och fermentationen uppströms, som ger substrat till metanogenerna.
Methanoregula är allestädes närvarande och mycket riklig i alla fyra grupperna av terrestra livsmiljöer (figur 6), men praktiskt taget obefintlig i det marina systemet, och kan därför visa sig vara en proxy för sötvattensinflytande i den marina sfären. Dess globala relevans rapporterades nyligen på annat håll (Yang et al., 2017). Trots sin allestädes närvarande spridning i jordar och sjösediment förekommer Methanoregula som en indikatorlinje i sura livsmiljöer med måttliga temperaturer. Dessutom verkar (1) Methanolinea särskilt ha anpassat sig till varma, neutrala terrestra miljöer, (2) Methanobacterium till kalla, sura miljöer och (3) Methanosaeta till pH-neutrala miljöer, vilket stämmer överens med andra studier (Rosenberg et al., 2014) och understryker robustheten i vårt tillvägagångssätt. Generellt sett kommer de geokemiska förhållanden som omger metanogena samhällen att leda till nischdifferentiering. Eftersom nischsorteringen tenderar att lämna de adaptiva specialisterna (Langenheder och Székely, 2011), genererade det progressiva långsiktiga miljöurvalet en mängd olika nischer som fylldes av en rad endemiska habitatspecialister, som kan vara mindre representerade eller frånvarande i andra olika miljöförhållanden. Samhället formas också av biotiska faktorer, t.ex. ekologiska interaktioner, dynamik, konkurrens och symbios. Trots dessa biologiska faktorer fann von Mering et al. (2007) att habitatpreferenser ofta är anmärkningsvärt stabila över tid och att den distinkta taxonomiska sammansättningen av miljösamhällen i sin tur kan vara en viktig indikator på deras ekologi och funktion.
Sammanhängande med habitatpreferenserna hos metanogena arkéer verkar det som om närbesläktade metanogena stammar ofta isolerades från jämförbara miljöer. Till exempel verkar Methanoregulaceae vara ganska varierande i naturliga miljöer (Yang et al., 2017) och neutrala stammar kan motstå odling hittills. De för närvarande beskrivna stammarna av Methanoregula har båda erhållits från svagt acidofila miljöer, medan de två representanterna av Methanolinea kommer från relativt varma livsmiljöer som rötkammarslam respektive risfältsjord (Rosenberg et al., 2014). Även om Methanobacterium-stammar isolerades från olika miljöer uppvisar ungefär hälften av de befintliga isolaten av detta släkte pH-optima som är något lägre än 7. Detta innebär att de indikatorlinjestammar som identifierades baserat på miljösekvenserna skulle kunna återspegla differentieringarna av fysiologi och källor hos de befintliga metanogena odlingarna. Till exempel kan salthalten i livsmiljön, som en allmän egenskap hos livsmiljön, successivt utsätta organismer för ett starkt miljömässigt urval och filtrera sammansättningen av en ny uppsättning arter som är bäst lämpade för den omgivande salthalten (Logares et al., 2013).
Bevarande och förvaltning av den biologiska mångfalden är en av de främsta utmaningarna i vårt nuvarande samhälle. Här visar vi att metanogena arkéer i naturliga miljöer är mest diversifierade i sediment från flodmynningar. Estuarier är övergångszoner mellan marina och terrestra ekosystem. Detta möjliggör två viktiga processer som kan bidra till artrikedomen. För det första blandas mikrober från hav och land i flodmynningarna och omfattar så småningom en övergripande hög mångfald (McLusky och Elliott, 2004). Man har till exempel observerat den höga mångfalden av bakterier, arkéer, svampar och till och med specifika bakterier som utför unika funktioner i flodmynningar (Cunliffe et al., 2008; Mosier och Francis, 2008; Crump et al., 2012). En annan aspekt är den höga näringsnivån på grund av de terrestriska, tidvatteninflödena som estuariernas organismer kan livnära sig på (McLusky och Elliott, 2004; Statham, 2012). I detta sammanhang är estuariemiljöerna viktiga för att återskapa generisk nyhet för metanogener. Hittills har effekterna av artdiversitet på ekosystemprocesser lockat till sig betydande forskningsinsatser. Kopplingen mellan biologisk mångfald och ekosystemfunktion diskuteras fortfarande och förblir svårdefinierad för mikrobiella samhällen (Loreau et al., 2001; Tilman et al., 2014). Även om jordar och sjösediment är primära källor till metan och även livsmiljöer med hög metanogen mångfald, föreslår vi att artrikedom inte är en lämplig proxy för metanproduktionspotential och ekosystemets metanutsläpp; den verkar snarare återspegla miljöns heterogenitet och historia. Att rangordna miljöerna enligt deras artrikedom innebär inte nödvändigtvis potentialen för metanutsläpp som är högst från jordar och sjöar och jämförelsevis mindre från flodmynningar (figur 3 och kompletterande figur S1).
För det sista kan bristen på miljöinformation i de offentliga databaserna ha försvårat en fullständig tolkning av de miljömässiga drivkrafter som observerats här. Även om det finns allt fler sekvenseringsdata om metanogener i litteraturen och offentliga databaser, är de relaterade abiotiska variabler som tillhandahålls ofta inkonsekventa och sparsamma. Den begränsade mängden konsekvent information om miljövariabler begränsar tillämpningen av statistiska analyser med flera variabler. Om man tänker på att de abiotiska faktorerna i den här studien endast kan förklara en begränsad del av variationerna i samhället tyder det på att det saknas andra förklarande variabler. Särskilt viktigt kan vara koncentrationerna och tillgången på metanogena substrat som acetat, väte och metylaminer. Icke desto mindre kan abiotiska parametrar aldrig vara tillräckliga för att fullt ut förklara vad som strukturerar metanogena sammansättningar, helt enkelt därför att livsmiljöer har olika historik och endast kan studeras lokalt. Bland de tillgängliga mcrA-uppgifterna är de stora områdena i den ryska och kanadensiska subarktiska och arktiska regionen dåligt representerade. En bättre geografisk täckning och jämnare fördelning av mcrA-genuppsättningen skulle förbättra bedömningen av metanogena samhällen på global nivå.
Författarnas bidrag
SL och XW utformade studien. XW och SY samlade in och analyserade data. XW, SY och FH utförde den statistiska analysen. XW, MW och SY gjorde den fylogenetiska korrigeringen. XW, SY, FH, MW, DW och SL tolkade resultaten och skrev artikeln. Alla författare bidrog till diskussionerna och granskade manuskriptet.
Intressekonfliktförklaring
Författarna förklarar att forskningen utfördes i avsaknad av kommersiella eller ekonomiska relationer som skulle kunna tolkas som en potentiell intressekonflikt.
Acknowledgments
Det ekonomiska stödet till XW (Grant No. 201408620031 till XW) som tillhandahölls av China Scholarship Council (CSC) är tacksamt erkänt. Denna studie stöddes av Helmholtz Gemeinschaft (HGF) genom finansiering av Helmholtz Young Investigators Group of SL (VH-NG-919).
Supplementary Material
Supplementary Material for this article can be found online at: https://www.frontiersin.org/article/10.3389/fmicb.2017.01339/full#supplementary-material
Fotnoter
- ^ http://metanogen.biotech.uni.wroc.pl/
- ^ http://www.ncbi.nlm.nih.gov/
- ^ http://metanogen.biotech.uni.wroc.pl/
- ^ https://www.ngdc.noaa.gov/mgg/shorelines/gshhs.html
- ^ http://qgis.osgeo.org
Auguet, J. C., Barberan, A. och Casamayor, E. O. (2010). Globala ekologiska mönster hos okultiverade archaea. ISME J. 4, 182-190. doi: 10.1038/ismej.2009.109
PubMed Abstract | CrossRef Full Text | Google Scholar
Bange, H. W., Bartell, U. H., Rapsomanikis, S. och Andreae, M. O. (1994). Metan i Östersjön och Nordsjön och en ny bedömning av de marina utsläppen av metan. Glob. Biogeochem. Cycles 8, 465-480. doi: 10.1029/94GB02181
CrossRef Full Text | Google Scholar
Barreto, D. P., Conrad, R., Klose, M., Claus, P. och Enrich-Prast, A. (2014). Avståndsförfall och taxa-area-relationer för bakterier, arkéer och metanogena arkéer i ett tropiskt sjösediment. PLoS ONE 9:e110128. doi: 10.1371/journal.pone.0110128
PubMed Abstract | CrossRef Full Text | Google Scholar
Bartlett, K. B., Bartlett, D. S., Harriss, R. C. och Sebacher, D. I. (1987). Metanutsläpp längs en salthaltsgradient i saltmarker. Biogeochemistry 4, 183-202. doi: 10.1007/BF02187365
CrossRef Full Text | Google Scholar
Borrel, G., O’Toole, P. W., Harris, H. M. B., Peyret, P., Brugere, J. F. och Gribaldo, S. (2013). Fylogenomiska data stöder en sjunde ordning av metylotrofa metanogener och ger insikter i metanogenesens evolution. Genome Biol. Evol. 5, 1769-1780. doi: 10.1093/Gbe/Evt128
PubMed Abstract | CrossRef Full Text | Google Scholar
Bousquet, P., Ciais, P., Miller, J., Dlugokencky, E., Hauglustaine, D., Prigent, C., et al. (2006). Antropogena och naturliga källors bidrag till atmosfärisk metanvariabilitet. Nature 443, 439-443. doi: 10.1038/nature05132
PubMed Abstract | CrossRef Full Text | Google Scholar
Bridgham, S. D., Cadillo-Quiroz, H., Keller, J. K., and Zhuang, Q. (2013). Metanutsläpp från våtmarker: biogeokemiska, mikrobiella och modelleringsperspektiv från lokal till global skala. Glob. Chang. Biol. 19, 1325-1346. doi: 10.1111/gcb.12131
PubMed Abstract | CrossRef Full Text | Google Scholar
Cao, H., Auguet, J.-C. och Gu, J.-D. (2013). Globalt ekologiskt mönster för ammoniakoxiderande arkéer. PLoS ONE 8:e52853. doi: 10.1371/journal.pone.0052853
PubMed Abstract | CrossRef Full Text | Google Scholar
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Lozupone, C. A., Turnbaugh, P. J., et al. (2011). Globala mönster av 16S rRNA-diversitet på ett djup av miljontals sekvenser per prov. Proc. Natl. Acad. Sci. U.S.A. 108, 4516-4522. doi: 10.1073/pnas.1000080107
PubMed Abstract | CrossRef Full Text | Google Scholar
Chaffron, S., Rehrauer, H., Pernthaler, J. och von Mering, C. (2010). Ett globalt nätverk av samexisterande mikrober från miljö- och helgenomsekvensdata. Genome Res. 20, 947-959. doi: 10.1101/gr.104521.109
PubMed Abstract | CrossRef Full Text | Google Scholar
Church, M. J., Bjorkman, K. M., Karl, D. M., Saito, M. A. och Zehr, J. P. (2008). Regional fördelning av kvävefixerande bakterier i Stilla havet. Limnol. Oceanogr. 53, 63-77. doi: 10.4319/lo.2008.53.1.0063
CrossRef Full Text | Google Scholar
Conrad, R. (2007). ”Microbial ecology of methanogens and methanotrophs”, i Advances in Agronomy, ed. L. S. Donald (Cambridge, MA: Academic Press), 1-63.
Google Scholar
Cornwell, W. K., Schwilk, D. W. och Ackerly, D. D. (2006). Ett egenskapsbaserat test för habitatfiltrering: konvex skrovvolym. Ecology 87, 1465-1471. doi: 10.1890/0012-9658(2006)872.0.CO;2
PubMed Abstract | CrossRef Full Text | Google Scholar
Crump, B. C., Amaral-Zettler, L. A. och Kling, G. W. (2012). Den mikrobiella mångfalden i arktiska sötvatten struktureras av inokulering av mikrober från jordar. ISME J. 6, 1629-1639. doi: 10.1038/ismej.2012.9
PubMed Abstract | CrossRef Full Text | Google Scholar
Cui, M., Ma, A., Qi, H., Zhuang, X., Zhuang, G. och Zhao, G. (2015). Varmare temperatur påskyndar metanutsläpp från våtmarken Zoige på den tibetanska högplatån utan att förändra det metanogena samhällets sammansättning. Sci. Rep. 5:11616. doi: 10.1038/srep11616
PubMed Abstract | CrossRef Full Text | Google Scholar
Cunliffe, M., Schafer, H., Harrison, E., Cleave, S., Upstill-Goddard, R. och Murrell, J. C. (2008). Fylogenetisk och funktionell genanalys av bakterie- och arkealsamhällen som är associerade med ytmikroskiktet i en flodmynning. ISME J. 2, 776-789. doi: 10.1038/ismej.2008.28
PubMed Abstract | CrossRef Full Text | Google Scholar
Dray, S., and Dufour, A. B. (2007). Paketet ade4: implementering av dualitetsdiagrammet för ekologer. J. Stat. Softw. 22, 1-20. doi: 10.18637/jss.v022.i04
CrossRef Full Text | Google Scholar
Dufrene, M. och Legendre, P. (1997). Artsammansättningar och indikatorarter: behovet av ett flexibelt asymmetriskt tillvägagångssätt. Ecol. Monogr. 67, 345-366. doi: 10.2307/2963459
CrossRef Full Text | Google Scholar
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C. och Knight, R. (2011). UCHIME förbättrar känsligheten och snabbheten vid upptäckt av chimärer. Bioinformatics 27, 2194-2200. doi: 10.1093/bioinformatics/btr381
PubMed Abstract | CrossRef Full Text | Google Scholar
Evans, P. N., Parks, D. H., Chadwick, G. L., Robbins, S. J., Orphan, V. J., Golding, S. D., et al. (2015). Metanmetabolism i archaeal phylum Bathyarchaeota avslöjad av genomcentrerad metagenomik. Science 350, 434-438. doi: 10.1126/science.aac7745
PubMed Abstract | CrossRef Full Text | Google Scholar
Fish, J. A., Chai, B., Wang, Q., Sun, Y., Brown, C. T., Tiedje, J. M., et al. (2013). FunGene: den funktionella genpipeline och det funktionella genregistret. Front. Microbiol. 4:291. doi: 10.3389/fmicb.2013.00291
PubMed Abstract | CrossRef Full Text | Google Scholar
Frank-Fahle, B. A., Yergeau, E., Greer, C. W., Lantuit, H. och Wagner, D. (2014). Mikrobiell funktionell potential och samhällssammansättning i permafrostpåverkade jordar i nordvästra kanadensiska Arktis. PLoS ONE 9:e84761. doi: 10.1371/journal.pone.0084761.g001
PubMed Abstract | CrossRef Full Text | Google Scholar
Fukuzaki, S., Nishio, N. och Nagai, S. (1990). Kinetik för metanogen jäsning av acetat. Appl. Environ. Microbiol. 56, 3158-3163.
Google Scholar
Hanson, C. A., Fuhrman, J. A., Horner-Devine, M. C. och Martiny, J. B. (2012). Bortom biogeografiska mönster: processer som formar det mikrobiella landskapet. Nat. Rev. Microbiol. 10, 497-506. doi: 10.1038/Nrmicro2795
PubMed Abstract | CrossRef Full Text | Google Scholar
Holm, G. O., Perez, B. C., McWhorter, D. E., Krauss, K. W., Johnson, D. J., Raynie, R. C., et al. (2016). Metanflöden på ekosystemnivå från tidvattens sötvatten- och bräcktkärr i Mississippiflodens delta: konsekvenser för kolprojekt för våtmarker vid kusten. Wetlands 36, 401-413. doi: 10.1007/s13157-016-0746-7
CrossRef Full Text | Google Scholar
Hunger, S., Schmidt, O., Hilgarth, M., Horn, M. A., Kolb, S., Conrad, R., et al. (2011). Konkurrerande format- och koldioxidförbrukande prokaryoter i en anoxisk metanavgivande mossjord. Appl. Environ. Microbiol. 77, 3773-3785. doi: 10.1128/AEM.00282-11
PubMed Abstract | CrossRef Full Text | Google Scholar
IPCC (2007). ”Climate Change 2007: the Physical Science Basis”, i Proceedings of the Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, eds D. Qin, S. Solomon, M. Manning, Z. Chen, M. Marquis, K. B. Averyt, et al. (Cambridge: Cambridge University Press).
Google Scholar
Jabłoński, S., Rodowicz, P. och Łukaszewicz, M. (2015). Databas över metanogena arkéer som innehåller fysiologiska och biokemiska egenskaper. Int. J. Syst. Evol. Microbiol. 65, 1360-1368. doi: 10.1099/ijs.0.000065
PubMed Abstract | CrossRef Full Text | Google Scholar
Kirschke, S., Bousquet, P., Ciais, P., Saunois, M., Canadell, J. G., Dlugokencky, E. J., et al. (2013). Tre decennier av globala metankällor och sänkor. Nat. Geosci. 6, 813-823. doi: 10.1038/Ngeo1955
CrossRef Full Text | Google Scholar
Knief, C. (2015). Mångfald och habitatpreferenser hos odlade och okultiverade aeroba metanotrofa bakterier utvärderade utifrån pmoA som molekylär markör. Front. Microbiol. 6:1346. doi: 10.3389/fmicb.2015.01346
PubMed Abstract | CrossRef Full Text | Google Scholar
Kotsyurbenko, O., Friedrich, M., Simankova, M., Nozhevnikova, A., Golyshin, P., Timmis, K., et al. (2007). Förändring från acetoklastisk till H2-beroende metanogenes i en västsibirisk torvmosse vid låga pH-värden och isolering av en acidofil Methanobacterium-stam. Appl. Environ. Microbiol. 73, 2344-2348. doi: 10.1128/AEM.02413-06
PubMed Abstract | CrossRef Full Text | Google Scholar
Langenheder, S., and Székely, A. J. (2011). Artssortering och neutrala processer är båda viktiga under den inledande sammansättningen av bakteriesamhällen. ISME J. 5, 1086-1094. doi: 10.1038/ismej.2010.207
PubMed Abstract | CrossRef Full Text | Google Scholar
Liebner, S., Ganzert, L., Kiss, A., Yang, S., Wagner, D. och Svenning, M. M. (2015). Skiftningar i metanogena samhällets sammansättning och metanflöden längs nedbrytningen av diskontinuerlig permafrost. Front. Microbiol. 6:356. doi: 10.3389/fmicb.2015.00356
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, Y. Q., Priscu, J. C., Xiong, J. B., Conrad, R., Vick-Majors, T., Chu, H. Y., et al. (2016). Salthalt driver arkealiska spridningsmönster i höghöjdssediment från sjöar på den tibetanska högplatån. FEMS Microbiol. Ecol. 92:fiw033. doi: 10.1093/femsec/fiw033
PubMed Abstract | CrossRef Full Text | Google Scholar
Logares, R., Lindstrom, E. S., Langenheder, S., Logue, J. B., Paterson, H., Laybourn-Parry, J., et al. (2013). Biogeografi hos bakteriesamhällen som utsätts för progressiva långsiktiga miljöförändringar. ISME J. 7, 937-948. doi: 10.1038/ismej.2012.168
PubMed Abstract | CrossRef Full Text | Google Scholar
Loreau, M., Naeem, S., Inchausti, P., Bengtsson, J., Grime, J. P., Hector, A., et al. (2001). Biologisk mångfald och ekosystemens funktion: nuvarande kunskap och framtida utmaningar. Science 294, 804-808. doi: 10.1126/science.1064088
PubMed Abstract | CrossRef Full Text | Google Scholar
Lozupone, C. A., and Knight, R. (2007). Globala mönster i den bakteriella mångfalden. Proc. Natl. Acad. Sci. U.S.A. 104, 11436-11440. doi: 10.1073/pnas.0611525104
PubMed Abstract | CrossRef Full Text | Google Scholar
Luton, P. E., Wayne, J. M., Sharp, R. J. och Riley, P. W. (2002). McrA-genen som ett alternativ till 16S rRNA i den fylogenetiska analysen av metanogenpopulationer i deponier. Microbiology 148, 3521-3530. doi: 10.1099/00221287-148-11-3521
PubMed Abstract | CrossRef Full Text | Google Scholar
Lutz, S., Anesio, A. M., Raiswell, R., Edwards, A., Newton, R. J., Gill, F., et al. (2016). Biogeografi av mikrobiomer i röd snö och deras roll i smältande arktiska glaciärer. Nat. Commun. 7:11968. doi: 10.1038/ncomms11968
PubMed Abstract | CrossRef Full Text | Google Scholar
Martiny, J. B. H., Bohannan, B. J. M., Brown, J. H., Colwell, R. K., Fuhrman, J. A., Green, J. L., et al. (2006). Mikrobiell biogeografi: att sätta mikroorganismer på kartan. Nat. Rev. Microbiol. 4, 102-112. doi: 10.1038/Nrmicro1341
PubMed Abstract | CrossRef Full Text | Google Scholar
Martiny, J. B. H., Eisen, J. A., Penn, K., Allison, S. D. och Horner-Devine, M. C. (2011). Drivers of bacterial beta-diversity depend on spatial scale. Proc. Natl. Acad. Sci. U.S.A. 108, 7850-7854. doi: 10.1073/pnas.1016308108
PubMed Abstract | CrossRef Full Text | Google Scholar
McArdle, B. H., and Anderson, M. J. (2001). Anpassning av multivariata modeller till samhällsdata: en kommentar till distansbaserad redundansanalys. Ecology 82, 290-297. doi: 10.1890/0012-9658(2001)0822.0.CO;2
CrossRef Full Text | Google Scholar
McCalley, C. K., Woodcroft, B. J., Hodgkins, S. B., Wehr, R. A., Kim, E.-H., Mondav, R., et al. (2014). Metandynamik som regleras av mikrobiella samhällen som reagerar på permafrostens upptining. Nature 514, 478-481. doi: 10.1038/nature13798
PubMed Abstract | CrossRef Full Text | Google Scholar
McGenity, T. J. (2010). ”Methanogens and Methanogenesis in Hypersaline Environments”, i Handbook of Hydrocarbon and Lipid Microbiology, red. K. N. Timmis (Berlin: Springer), 665-680. doi: 10.1007/978-3-540-77587-4_53
CrossRef Full Text | Google Scholar
McLusky, D. S., and Elliott, M. (2004). Det estuarina ekosystemet: Ecology, Threats and Management (Ekologi, hot och förvaltning). New York, NY: Oxford University Press. doi: 10.1093/acprof:oso/9780198525080.001.0001
CrossRef Full Text | Google Scholar
Megonigal, J. P., Mines, M. E., and Visscher, P. T. (2005). ”Anaerobic metabolism: linkages to trace gases and aerobic processes”, i Biogeochemistry, ed. W. H. Schlesinger (Oxford: Elsevier), 350-362.
Google Scholar
Milferstedt, K., Youngblut, N. D. och Whitaker, R. J. (2010). Rumslig struktur och persistens hos metanogenpopulationer i humösa myrsjöar. ISME J. 4, 764-776. doi: 10.1038/Ismej.2010.7
PubMed Abstract | CrossRef Full Text | Google Scholar
Mosier, A. C., and Francis, C. A. (2008). Relativ abundans och mångfald av ammoniakoxiderande archaea och bakterier i San Francisco Bay estuary. Environ. Microbiol. 10, 3002-3016. doi: 10.1111/j.1462-2920.2008.01764.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Nazaries, L., Murrell, J. C., Millard, P., Baggs, L., and Singh, B. K. (2013). Metan, mikrober och modeller: grundläggande förståelse av markens metancykel för framtida prognoser. Environ. Microbiol. 15, 2395-2417. doi: 10.1111/1462-2920.12149
PubMed Abstract | CrossRef Full Text | Google Scholar
Nelson, M. B., Martiny, A. C., and Martiny, J. B. H. (2016). Global biogeografi av mikrobiella egenskaper för kvävecykling i marken. Proc. Natl. Acad. Sci. U.S.A. 113, 8033-8040. doi: 10.1073/pnas.1601070113
PubMed Abstract | CrossRef Full Text | Google Scholar
Nemergut, D. R., Costello, E. K., Hamady, M., Lozupone, C., Jiang, L., Schmidt, S. K., et al. (2011). Globala mönster i biogeografin hos bakterietaxa. Environ. Microbiol. 13, 135-144. doi: 10.1111/j.1462-2920.2010.02315.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Nobu, M. K., Narihiro, T., Kuroda, K., Mei, R. och Liu, W. T. (2016). Chasing the elusive Euryarchaeota class WSA2: genomes reveal a uniquely fastidious methyl-reducing methanogen. ISME J. 10, 2478-2487. doi: 10.1038/ismej.2016.33
PubMed Abstract | CrossRef Full Text | Google Scholar
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O’Hara, R. B., et al. (2015). vegan: Community Ecology Package. R Package Version 2.2.0. Tillgänglig på: https://CRAN.R-project.org/package=vegan
Google Scholar
Oloo, F., Valverde, A., Quiroga, M. V., Vikram, S., Cowan, D. och Mataloni, G. (2016). Habitatets heterogenitet och konnektivitet formar mikrobiella samhällen i sydamerikanska torvmarker. Sci. Rep. 6:25712. doi: 10.1038/srep25712
PubMed Abstract | CrossRef Full Text | Google Scholar
Pattnaik, P., Mishra, S. R., Bharati, K., Mohanty, S. R., Sethunathan, N. och Adhya, T. K. (2000). Inverkan av salthalt på metanogenes och tillhörande mikroflora i tropiska risjordar. Microbiol. Res. 155, 215-220. doi: 10.1016/S0944-5013(00)80035-X
PubMed Abstract | CrossRef Full Text | Google Scholar
Poffenbarger, H. J., Needelman, B. A., and Megonigal, J. P. (2011). Salinitetens inverkan på metanutsläpp från tidvattensmarker. Wetlands 31, 831-842. doi: 10.1007/s13157-011-0197-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Potter, E. G., Bebout, B. M., and Kelley, C. A. (2009). Isotopisk sammansättning av metan och metanogena substrat längs en salthaltsgradient i ett hypersalt mikrobiellt mattsystem. Astrobiology 9, 383-390. doi: 10.1089/ast.2008.0260
PubMed Abstract | CrossRef Full Text | Google Scholar
R Core Team (2014). R: A Language and Environment for Statistical Computing (Version 3.0.3). Wien: R Foundation for Statistical Computing.
Google Scholar
Roberts, D. (2016). labdsv: Ordination och multivariat analys för ekologi. R Package Version 1.8-0. Tillgänglig på: https://CRAN.R-project.org/package=labdsv
Google Scholar
Rooney-Varga, J. N., Giewat, M. W., Duddleston, K. N., Chanton, J. P. och Hines, M. E. (2007). Kopplingar mellan arkeologiska samhällsstrukturer, vegetationstyp och metanogena vägar i torvmarker i Alaska. FEMS Microbiol. Ecol. 60, 240-251. doi: 10.1111/j.1574-6941.2007.00278.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Rosenberg, E., DeLong, E. F., Lory, S., Stackebrandt, E. och Thompson, F. (2014). The Prokaryotes: Andra större linjer av bakterier och arkéer. Berlin: Springer.
Google Scholar
Ruff, S. E., Biddle, J. F., Teske, A. P., Knittel, K., Boetius, A. och Ramette, A. (2015). Global spridning och lokal diversifiering av mikrobiomet i metanutsläpp. Proc. Natl. Acad. Sci. U.S.A. 112, 4015-4020. doi: 10.1073/pnas.1421865112
PubMed Abstract | CrossRef Full Text | Google Scholar
Schauer, R., Bienhold, C., Ramette, A. och Harder, J. (2009). Bakteriediversitet och biogeografi i djuphavssediment på ytan i södra Atlanten. ISME J. 4, 159-170. doi: 10.1038/ismej.2009.106
PubMed Abstract | CrossRef Full Text | Google Scholar
Schloss, P. D., Westcott, S. L., Ryabin, T., Hall, J. R., Hartmann, M., Hollister, E. B., et al. Introducing mothur: öppen källkod, plattformsoberoende, gemenskapsstödd programvara för att beskriva och jämföra mikrobiella samhällen. Appl. Environ. Microbiol. 75, 7537-7541. doi: 10.1128/Aem.01541-09
PubMed Abstract | CrossRef Full Text | Google Scholar
Statham, P. J. (2012). Näringsämnen i flodmynningar – en översikt och klimatförändringens potentiella effekter. Sci. Total Environ. 434, 213-227. doi: 10.1016/j.scitotenv.2011.09.088
PubMed Abstract | CrossRef Full Text | Google Scholar
Thauer, R. K., Kaster, A.-K., Seedorf, H., Buckel, W. och Hedderich, R. (2008). Metanogena arkéer: ekologiskt relevanta skillnader i energihushållning. Nat. Rev. Microbiol. 6, 579-591. doi: 10.1038/Nrmicro1931
PubMed Abstract | CrossRef Full Text | Google Scholar
Tilman, D., Isbell, F., and Cowles, J. M. (2014). Biologisk mångfald och ekosystemens funktion. Annu. Rev. Ecol. Evol. Syst. 45, 471-493. doi: 10.1146/annurev-ecolsys-120213-091917
CrossRef Full Text | Google Scholar
Ulrich, W., Hajdamowicz, I., Zalewski, M., Stañska, M., Ciurzycki, W. och Tykarski, P. (2010). Artutbud eller filtrering av livsmiljöer: en fallstudie av spindelsamhällen på öar i en sjö. Ecol. Res. 25, 375-381. doi: 10.1007/s11284-009-0661-y
CrossRef Full Text | Google Scholar
Vanwonterghem, I., Evans, P. N., Parks, D. H., Jensen, P. D., Woodcroft, B. J., Hugenholtz, P., et al. (2016). Metylotrofisk metanogenes upptäckt i den arkeala fylummet Verstraetearchaeota. Nat. Microbiol. 1:16170. doi: 10.1038/nmicrobiol.2016.170
PubMed Abstract | CrossRef Full Text | Google Scholar
von Mering, C., Hugenholtz, P., Raes, J., Tringe, S. G., Doerks, T., Jensen, L. J., et al. (2007). Kvantitativ fylogenetisk bedömning av mikrobiella samhällen i olika miljöer. Science 315, 1126-1130. doi: 10.1126/science.1133420
PubMed Abstract | CrossRef Full Text | Google Scholar
Wagner, D., and Liebner, S. (2009). ”Global warming and carbon dynamics in permafrost soils: methane production and oxidation”, i Permafrost Soils, red. R. Margesin (Berlin: Springer), 219-236.
Google Scholar
Weiher, E., and Keddy, P. (2001). Ekologiska församlingsregler: Perspektiv, framsteg, reträtter. Cambridge: Cambridge University Press.
Google Scholar
Whitaker, R. J., Grogan, D. W. och Taylor, J. W. (2003). Geografiska barriärer isolerar endemiska populationer av hypertermofila archaea. Science 301, 976-978. doi: 10.1126/science.1086909
PubMed Abstract | CrossRef Full Text | Google Scholar
Wickham, H. (2009). ggplot2: Elegant Graphics for Data Analysis. New York, NY: Springer-Verlag. doi: 10.1007/978-0-387-98141-3
CrossRef Full Text | Google Scholar
Yang, J., Ma, L. A., Jiang, H., Wu, G. och Dong, H. (2016). Salthalt formar mikrobiell mångfald och samhällsstruktur i ytsediment i Qinghai-Tibetanska sjöarna. Sci. Rep. 6:25078. doi: 10.1038/srep25078
PubMed Abstract | CrossRef Full Text | Google Scholar
Yang, S., Liebner, S., Alawi, M., Ebenhöh, O. och Wagner, D. (2014). Taxonomisk databas och gränsvärde för bearbetning av mcrA-genens 454-pyrosekvenseringsdata med MOTHUR. J. Microbiol. Methods 103, 3-5. doi: 10.1016/j.mimet.2014.05.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Yang, S., Liebner, S., Winkel, M., Alawi, M., Horn, F., Dörfer, C., et al. (2017). Djupgående analys av metanogena samhällen i kärnor från permafrostpåverkade våtmarker på hög höjd. Soil Biol. Biochem. 111, 66-77. doi: 10.1016/j.soilbio.2017.03.007
CrossRef Full Text | Google Scholar
Ye, R. Z., Jin, Q. S., Bohannan, B., Keller, J. K., McAllister, S. A., and Bridgham, S. D. (2012). pH-kontroller över anaerob kolmineralisering, metanproduktionens effektivitet och metanogena vägar i torvmarker över en ombrotrofisk-minerotrofisk gradient. Soil Biol. Biochem. 54, 36-47. doi: 10.1016/j.soilbio.2012.05.015
CrossRef Full Text | Google Scholar
Zinder, S. H. (1993). ”Physiological ecology of methanogens”, i Methanogenesis: Ecology, Physiology, Biochemistry & Genetics, ed. J. G. Ferry (Boston, MA: Springer), 128-206.
Google Scholar
Zinger, L., Amaral-Zettler, L. A., Fuhrman, J. A., Horner-Devine, M. C., Huse, S. M., Welch, D. B. M., et al. (2011). Globala mönster av bakteriell beta-diversitet i ekosystem på havsbotten och i havsvatten. PLoS ONE 6:e24570. doi: 10.1371/journal.pone.0024570
PubMed Abstract | CrossRef Full Text | Google Scholar